Contents
Summary
Zusammenfassung
1 Introduction
1.1 Distribution and utilisation of setts
1.2 Distribution and utilisation of latrines
1.3 Specific questions
2 The "Sihlwald" study area
3 Methods
3.1 Collection of data
3.2 Analysis
4 Distribution and habitat analysis of setts
4.1 Size, distribution and density of setts
4.2 Type of sett entrances
4.3 Habitat analysis of setts
4.4 The habitat parameters
4.5 Discussion of the sett distribution, density and types of entrances
4.6 Discussion and synthesis of the habitat characteristics of the setts
5 Distribution and habitat analysis of latrines
5.1 Size, utilisation, distribution and density of latrines
5.2 Latrine location
5.3 Habitat analysis of latrines
5.4 Discussion of the distribution, density and utilisation of latrines
5.5 Discussion of the habitat analysis of latrines
6 Sihlwald - an ideal badger habitat
7 References
Summary
Certain species, like the badger (Meles meles), adapt their social and spatial organisation to different environments. Knowledge about the distribution of the setts and latrines may help to determine whether badgers in Sihlwald live in groups, have distinct home ranges and occupy a main sett or whether they are solitary, have overlapping homeranges and frequent different setts within the range.
The goal of this study was:
- to gather information about the distribution and the density of the setts in Sihlwald;
- to find out which habitat parameters affect the distribution of setts and latrines (habitat analysis);
- to see if the distribution of the setts and latrines correlates with the availability of those parameters in the forest.
From March-August 1996, 123 setts and 40 latrines in Sihlwald were found by intensive searching of the study area. Certain habitat parameters were recorded for each sett and latrine; some of the parameters were measured in the field, others were obtained from the GIS which contained a digital terrain model of the forest. To measure the availability of the habitat parameters, random points were recorded in the same way as the setts and latrines. The distribution of the setts was tested against a random distribution (i.e. the distribution of the nearest neighbour distance between setts was compared with the distribution of the nearest neighbour distance between random points). Areas of higher and lower density than would be expected from a random distribution were calculated using a Bivariate Normal Kernel Density Estimation. The habitat analysis was calculated by using a stepwise logistic regression. The distribution of the setts and latrines in certain habitat parameters were compared with the availability of those parameters with a goodness-of-fit test for GIS data, and with a Chi
2 -test for two independent samples for the field data.
The observed distribution of the setts differs significantly from random. The high sett density in Sihlwald (12.3/100 ha) indicates an ideal badger habitat. Setts were especially aggregated from "Häuliboden" to "Sihlzopf". Ideal sett sites were found above 600 metres a.s.l., closer to the forest boundary and adjoining agricultural zones than the random points. Furthermore, setts in Sihlwald are dug on convex slopes with an inclination of 20°-40°. Setts were found more than 50 metres from the nearest path and in areas with a sparse ground cover. Coniferous stands were avoided. However, single spruces were frequently used as sett sites for setts consisting of 1-2 entrances only. The density of the latrines in Sihlwald is 4/100 ha. Latrines were found above 600 metres a.s.l. and closer to the forest boundary than random points. Two thirds of the latrines were found near a conspicuous landmark.
The size of setts in Sihlwald indicate possible differences in sett utilisation. The distribution of setts and latrines indicate that certain badgers leave the forest to forage for food, whereas data of the radio-collared badgers show thast certain individuals never leave the forest. This leads to the assumption that several strategies may exist side by side: from individuals that live in groups, have distinct home ranges and occupy a main sett, to individuals that are solitary, have large overlapping home ranges, forage outside the forest and occupy several setts within a range.
Zusammenfassung
In verschiedenen Gebieten durchgeführte Untersuchungen lassen annehmen, dass Dachse ihr Sozialverhalten und ihre Raumnutzung auf das lokale Ressourcenangebot abzustimmen vermögen. Ausgehend davon ging es bei der vorliegenden Feldstudie darum, das Raummuster und die Habitatparameter der Baue und Latrinen im Sihlwald festzustellen. Erwartet wurde, dass die Befunde einen Beitrag liefern zur Frage, ob die Dachse in diesem Gebiet in Gruppen leben, voneinander getrennte Streifgebiete besitzen und einen Hauptbau bewohnen oder ob sie Einzelgänger sind, überlappende Streifgebiete haben und mehrere Baue in ihrem Streifgebiet benutzen.
Der Zweck der vorliegende Studie war:
Die Feldarbeit wurde zwischen März und August 1996 ausgeführt. Dabei wurden 123 Baue und 40 Latrinen im Sihlwald mittels intensivem Absuchen des Untersuchungsgebietes gefunden. Für jeden Bau und jede Latrine wurden ausgewählte Habitatparameter aufgenommen. Ergänzend dazu, sind dem GIS, das ein digitales Geländemodel des Sihlwaldes enthält, Habitatparameter entnommen worden. Um das Angebot der Habitatparameter ermitteln zu können, sind an Zufallspunkten die gleichen Habitatparameter aufgenommen worden. Die räumliche Verteilung der Baue wurde mit einer zufälligen Verteilung verglichen (d.h. die Verteilung der Distanz zwischen den Bauen wurde mit jener zwischen Zufallspunkten verglichen). Mittels einer Bivariat-Normalen Kernel Dichteschätzung sind Flächen mit geringerer und höherer Dichte an Bauen und Latrinen ermittelt worden. Die Habitatsanalyse wurde mit einer schrittweisen Logistischen Regression ausgewertet. Die Verteilung der Baue und Latrinen in den GIS-Habitatparametern wurde mittels einem Chi
2-Anpassungstest mit dem Angebot verglichen. Die im Feld erhobenen Habitatparameter sind mit einem Chi
2-Test für zwei unabhängige Stichproben mit dem Angebot verglichen worden.
Die räumliche Verteilung der Baue unterscheidet sich signifikant von einer zufälligen räumlichen Anordnung. Die Dichte der Baue im Sihlwald (10 km2) beträgt 12.3/100 ha. Besonders zwischen "Häuliboden" und "Sihlzopf" sind die Baue auffällig gehäuft. Bevorzugte Standorte liegen über 600 m.ü.M. und befinden sich näher an Waldrändern mit anschliessendem Kulturland als die Zufallspunkte. Die Baue sind im Sihlwald vorzugsweise in konvexen Abhängen mit einer Neigung zwischen 20°-40° gegraben. Alle Baue liegen weiter als 50 Meter vom nächsten Weg entfernt in Gebieten mit spärlicher Bodenvegetation. Fichtenbestände werden gemieden. Bauen mit 1 oder 2 Eingängen werden jedoch häufig unter einzelnen Fichten angelegt. Die Dichte der Latrinen im Sihlwald beträgt 4/100 ha. Alle gefundenen Latrinen liegen über 600 m.ü.M. und näher am Waldrand als die Zufallspunkte. Zwei Drittel aller Latrinen befinden sich in der Nähe einer sichtbaren Landmarke.
Die Grösse der Baue im Sihlwald könnten einen Hinweis auf Unterschiede in der Art und Häufigkeit der Nutzung geben. Aufgrund der Verteilung der Baue und Latrinen scheint es, dass einzelne Individuen für die Nahrungssuche aus dem Wald gehen, wohingegen die radiotelemetrierten Dachse den Wald nie verlassen. Dies lässt vermuten, dass verschiedene Strategien nebeneinander vorkommen können: von Individuen, die in Gruppen leben, klar voneinander getrennte Streifgebiete besitzen und einen Hauptbau bewohnen, bis zu Individuen, die Einzelgänger sind, grosse sich überlappende Streifgebiete haben und mehrere Baue im Streifgebiet benutzen.
Acknowledgments
First of all I would like to thank Prof. B. Nievergelt for his friendship and for sharing his love for nature with me. The field trip to the Simen Mountains National Park in Ethiopia (Jan.-Feb. 1996) allowed me to get to know the people and projects of the department and to quickly feel at home here. My warmest thanks goes to Karin Hindenlang,who helped me organise my diploma and who supported me through all stages of it. In particular, I would like to express my gratitude to Stefan Imfeld, who generously helped me not only with all questions considering the GIS analysis, but with all other problems that arose during the statistical analysis of my data. I was extremely grateful for his critical remarks and moral support. Much thanks goes to Susanne Reimann for her flair for computers as well as for her talent as art director. Furthermore, I would like to thank Lorenz Gygax and Wolf Blankenhorn for the statistical advice and Gérald Achermann and Stefan Imfeld for helpful comments on the draft. Special thanks to André Utzinger for English corrections and the nice time at the hut. I would especially like to thank Mr. John Potts for proof-reading the manuscript.
I am grateful to many people who helped me find numerous setts: Gérald Achermann, Iris Bättig, Nina Debrunner, Oliver Fischer, Michi Griesser, Stefan Imfeld, Guido Keller, Isabelle Minder, Marco Muhmenthaler, Thomas Nabulon, Josef Niederberger, Dani Wirz and Matthias Wüst.
Introduction
Exploring the relationships between animal behaviour and ecology is very promising when a wide variability exists in a species’ social and spatial organisation: badgers (Meles meles) adapt their social and spatial organisation to different environments: in areas with a predictable food availability, badgers live in groups, defend territories and occupy a main sett; in areas with a wide seasonal variation in food availability, badgers are solitary, have large overlapping home ranges and use several setts within a range (Cheeseman et al., 1988; Cresswell & Harris, 1988; Skinner & Skinner, 1988). This extraordinary adaptability has undoubtedly contributed largely to their success as a species. Within the Swiss National Foundation grant 3100-40846.91 "Adjustment of spatial behaviour of badger (Meles meles L.) in response to a heterogeneous environment", a study on the European badger concentrates on how variation in resource availability between individual home ranges in the study area is reflected by variation in individual resource utilisation. In order to understand a part of the mechanisms of adjustment of the social and spatial organisation of badgers in different environments, this study focussed on the distribution and utilisation of setts and latrines in Sihlwald.
1.1 Distribution and utilisation of setts
Badgers are found all over Switzerland up to 2000 m a.s.l.. [ISS: figure omitted]. The badger has adapted to various habitats, but prefers the lowlands and hills. Its distribution in higher regions is limited by the absence of forests, fields and pastures (Graf, 1995). In contrast to the widespread and heterogeneous home ranges, the habitat requirements of the sett sites are rather specific.
As shown in the literature, the distribution and density of badger setts vary considerably between different countries and also between different locations within one country. There are many different factors affecting the distribution and density of badger setts, e.g. the type of soil, the amount of cover and the hilliness of the terrain (Neal, 1986). However, local conditions, particularly available food such as earthworms, also have an important modifying effect on sett choice (Skinner et al., 1991). Thus, the distribution of badgers is strongly determined by how well a certain area fulfils the following ecological requirements:
• a well-drained soil which is easy to dig and firm enough to prevent the roof of the sett from collapsing;
• sufficient cover to allow the animal to emerge and leave inconspicuously;
• a region in the immediate vicinity of the sett which is relatively free of disturbance from man or domestic animals;
• and the presence of an adequate food supply at all seasons of the year.
Setts vary greatly in size, the extent of the tunnels and the number of entrances. A number of them have been excavated and their structure compared (Roper, 1992). The size of a sett is not necessarily determined by the number of entrances, nor does sett size indicate the number of badgers living in it (Neal, 1977). Bock (1986) thinks that the volume of the spoil heaps is probably the best criterion to characterise and quantify a badger's sett.
Various attempts have been made to classify setts according to size and function (Kruuk, 1978; Thornton, 1988). In the studies carried out in Britain, the most important sett in a territory is termed "main sett" which is usually large, is used throughout the year and acts as the main breeding sett. In contrast, there are small setts with one or two holes and a simple, often unbranched tunnel system. They are used only occasionally as temporary refuges and are called "outliers". There may be several of these within a territory. Intermediate levels of size and complexity are found between these two extremes. Setts permanently in use may indicate territoriality; setts used only at certain times of the year (as winter refuge or in areas with a big seasonal variation in food availability) may point to a spatial organisation of extensive, overlapping home ranges.
Beside badgers, the most common sett tenants in Switzerland are foxes (Vulpes vulpes L.). Foxes occasionally dig their own den, but, being lazy diggers, they prefer badger setts when available (Weber, 1990). The fox is a surface dweller that requires secure cover below ground for rearing cubs. After the weaning of the cubs, setts are only frequented by foxes if the weather is harsh or very wet, or where surface cover is scarce (Weber, 1985). Typically, they use unoccupied setts, but in large setts share the spacious accommodation, usually using the perimeter holes (Neal & Cheeseman, 1996). For this reason, setts in Sihlwald were divided into the categories "used by badger", "used by a fox" and "used by badger and/or fox" (see Appendix, Tab. 13).
1.2 Distribution and utilisation of latrines
A latrine consists of one or more dung pits about 15 cm deep. When a pit is full of dung, it is left uncovered and another dug nearby. One can often find several dung pits covering an area of 2-4 m2. Occasionally, instead of a latrine with many pits, a single considerably larger one may be found containing much dung. Sometimes the dung is deposited on the surface, not in pits. Underground latrines found in excavated setts appear to be used when the badgers stay below in periods of very low temperature (Roper, 1992).
Kruuk (1978a) defined two types of latrines: those found near a sett and those that occur in other parts of the territory. The latrines near the border of ranges have territorial significance and are usually larger than those near setts. With regard to utilisation, Roper et al. (1986) distinguished two different types of defecation sites: latrines (several dung pits dug within an area of a few square metres and used more or less all year-round) and temporary defecation sites (one single dung pit in any one month; faeces found only during one or two months of the year). The former are usually located around territory boundaries, the latter elsewhere in the range. Some are near obvious landmarks such as hedgerows, fences or woodedges (Kruuk, 1978a; 1989; Roper et al., 1986).
1.3 Specific questions
Given the above-mentioned variability of the badger’s spatial and social organisation, knowledge about the distribution of setts and latrines may help to determine whether badgers in Sihlwald live in groups and occupy a main sett, or whether they are solitary and frequent different setts within the range. With this in mind, the following questions were examined for the habitat analysis of badger setts and latrines:
• How are the setts and latrines distributed in Sihlwald (randomly, clumped)?
• In what density do setts and latrines occur in this forest?
• Can the distribution of setts and/or latrines be explained by certain habitat parameters?
• Does the distribution of the setts and latrines correlate with the availability of the habitat parameters in Sihlwald?
2 The "Sihlwald" study area
This forest lies on the eastern flank of the Albis Range and runs from the SSE to the NNW between 8°31'-8°35'E and 47°13'-47°17'N. The elevation ranges from 470 m a.s.l. at the bottom of the valley to over 900 m a.s.l. on the ridge ([ISS: figure omitted]). Thus, Sihlwald finds itself between the submontane and montane zone and has the characteristic climate of the lower montane region (BGU, 1988) with an average annual rainfall of 1350 mm.
This forest has a pronounced topographical variation and is characterised by a diverse mosaic pattern of mixed forest, dominated by deciduous trees. Geologically speaking, it belongs to the Swiss plateau and consists of subalpine molassic sandstone with a partly morrainic cover. The dominant soil types are eutric cambisol and gleyic cambisol which contain mainly sandy to silty clay and argillaceous sand (Bodeneignungskarte der Schweiz 1: 50'000, 1980).
There are some very steep areas along the river Sihl and up on the ridge, but in between, at around 600 m a.s.l., gently sloping plateaux are furrowed by countless brooks. Sihlwald, recently declared a nature reserve, covers an area of approximately 1000 ha and is situated some 10 km south of the city of Zurich. This heterogeneous landscape has two ecological gradients: to the north and east a sharp gradient from forested to urban ranges; to the south and west a gradient from forested to agricultural zones.
3 Methods
3.1 Collection of data
The field work (March-August 1996) consisted in searching the study area all over for setts and latrines, using paths, brooks and forest divisions as borderlines. It is expected that most of, if not all, the existing setts were found. Owing to the often dense vegetation cover, it is assumed that only a small fraction of the latrines was actually discovered.
Sett definition
The definition of a badger sett in the Protection of Badgers Act (1992) in Britain is "any structure or place which displays signs indicating current use by a badger". Since this definition is rather vague, I defined a sett as at least one entrance leading more than two metres underground. Usually a large mound of excavated soil can be found outside each entrance. If two entrances were further than 25 m apart, they were noted as two separate setts. This definition is independent of the species using a particular sett, since in most areas no distinction could be made between setts used by badgers and those used by foxes. To analyse the distribution of the setts in Sihlwald, Kruuk’s (1978a) main sett definition (>2 entrances) was used to calculate the nearest neighbour distance between "main setts". Furthermore, five different sett types were distinguished: setts dug directly into the ground (Fig. 3), under a boulder/rock (Fig. 4), under a spruce (Picea abies) (Fig. 5), under a deciduous tree and under a tree stump.
Latrine definition
Latrines are aggregations of one to several defecations, deposited in one or in several small pits dug by the animals themselves. These dung pits are 5-10 cm deep, sometimes as much as 30 cm, and can contain several different defecations (Fig. 6). However, a few latrines have no pits, and some have all droppings in one large pit (Kruuk, 1978a). Since no distinction could be made in Sihlwald between empty dung pits and pits dug in search of food (beetles), a latrine had to contain dung in at least one pit to be defined as such.
Important habitat requirements as mentioned in chapter 1.1, as well as habitat parameters indirectly influencing sett and latrine sites, were recorded for each sett and latrine (see also Appendix , Tab. 12-14). To determine the availability of the recorded details, 86 random points were generated and recorded the same way as the setts and latrines. The coordinates of all setts, latrines and random points are given in the Appendix (Tab. 15-17). [ISS: This Appendix can only be viewed with the permission of the author (Password needed)]
Sett / latrine location: When a sett or latrine was found, its location was marked on the 1:10'000 map of Sihlwald. The coordinates were fed into a geographical information system, GIS, (Arc/Info V. 7.01) at the Institute for Geography at the University of Zurich for further analysis.
Altitude: The GIS contained a digital terrain model of Sihlwald with which the altitude was calculated.
Inclination: The inclination was recorded right at the sett or latrine site taking 10 m above and below the sett or latrine as measurement and put into the following categories for further analysis: 0-10°, 11-20°, 21-30°, 31-40°, >40°.
- Aspect: The aspect (compass direction of the slope) of the sett and latrine sites was calculated by the GIS and put into the following categories for further analysis: N (<=
45°), NE (<=
90°), E (<=
135°), SE (<=
180°), S (<=
225°), SW (<=
270°), W (<=
315°) and NW (<=
360°).
- Topography: The topography was taken within a radius of approximately 25 m of the centre of the sett or latrine. Six different types of topography were defined:
1. flat
2. gentle slope (neither convex nor concave)
3. concave slope
4. convex slope
5. crest
- Vegetation coverage: The degree of vegetation coverage (tree-, shrub-, herb- and moss coverage) around the sett was determined after Braun-blanquet (1964): 0=0%; 1=1-5%; 2=6-25%; 3=26-50%; 4=51-75%; 5= >75%.
- Forest parameters: The forest superintendent's office did a survey on upper-, middle- and lower layer coverage; upper-, middle- and lower coniferous layer coverage; vegetation unit and development stage of the forest stands (Stadtforstamt Zürich, 1990). This data can also be found in the GIS.
- Distance to the nearest path & Distance to the nearest water: These distances were calculated with the GIS and put into the following categories for further analysis: ¾ 50 m, ¾100 m, ¾150 m, ¾ 200 m, >200 m for distances to the nearest path; ¾ 50 m, ¾100 m, ¾ 150 m, >150 m for distances to the nearest water.
- Distance to forest boundary with adjoining agricultural zones (i.e. the forest boundary along the "Sihltalstrasse" was not included): These distances were calculated with the GIS and put into the following categories for further analysis: ¾ 100 m; ¾ 200 m; ....; ¾ 900 m; >900 m.
- Geological situation: Data on the geology of the region was taken from the digitised part of the 1:50'000 geology map of the Canton Zurich (R. Hantke, 1967).
Possible parameters affecting sett site are summarised in Fig. 7.The parameters printed in bold letters were chosen for this study. Species are affected by many things other than their habitat: parameters such as intra- and interspecific competition can have major effect on wildlife, meaning that a suitable habitat alone will not guarantee a species' survival.
3.2 Analysis
Analysis with the GIS
- Distribution: In order to test the distribution of the setts against a random distribution, the nearest neighbour distance between the setts was compared with the nearest neighbour distance for 1000 samples of 123 random points in the area.
- Density: To calculate areas with higher and lower sett density than expected from a random distribution, a Bivariate Normal Kernel Density Estimation was used in conjunction with a Monte Carlo simulation (Imfeld, in prep.).
Analysis with SPSS statistic package (V. 6.0) for Macintosh
- Habitat analysis: All parameters (GIS and field data) were plotted against each other to see if a linear correlation exists between them. Owing to the weak correlations, no parameter had to be omitted from further analysis.
In order to reduce the dimensionality of a set of factors, a principle components analysis (Mosteller & Tukey, 1977) was used. Because of the weak correlation between the parameters, no reduction of the parameters was attained.
For the habitat analysis of setts (i.e. to find out which of the 19 habitat parameters explain most of the variation for the distribution of the setts) a logistic regression estimated the coefficient of a probabilistic model, involving a set of independent parameters, that best predicts the value of a dichotomous variable (i.e. a variable that has two distinct values: "sett" and "no sett"). The parameters "geology", "vegetation unit" and "topography" were defined as categorical covariates (indicator contrast) equivalent to the traditional group of "dummy variables". The method chosen for the logistic regression was a backward stepwise selection. Removal testing was based on the probability of the likelihood ratio statistic based on the maximum partial likelihood estimates. Too few latrines were found to apply the logistic regression with the 19 parameters for the habitat analysis of latrines.
A residual analysis was then carried out to examine the possible violations of the normality assumptions (normal Q-Q plot of residuals; scatterplot of residuals versus the predicted values; scatterplot of residuals versus leverage; scatterplot of residuals versus individual independent variables).
- Distribution with regard to availability of habitat parameters: To test the random distribution of setts in the GIS habitat parameter categories (Appendix, Tab. 12), a goodness-of-fit test compared the number of observed setts with the number of expected setts for each category:

The number of setts was compared with the number of random points for each habitat parameter measured in the field (Field data, Appendix, Tab. 12) (Chi
2-test for two independent samples). The GIS and field data could not be analysed together, as the GIS parameters were available for the whole area of the forest, whereas the field data were not.
The habitat parameters for which the distribution of the setts was non-random were carefully analysed with a Chi
2-test in conjunction with a Bonferroni z statistic (Neu et al., 1974) to find out which parameter categories explain the distribution of the setts best. Trends were then tested with a Mann-Whitney U-test.
A Mann-Whitney U-test was applied in order to compare 1. the setts in areas with a higher sett density than expected from a random distribution (Bivariate Normal Kernel Density Estimation, see above) with all other setts, 2. the distance to forest boundary between setts and random points, and 3. the latrines near setts with those not in the vicinity of setts.
Small, middle-sized and big setts were compared with the Kruskal-Wallis 1-way Anova. With regard to the habitat parameters, the setts did not differ significantly from each other.
4 Distribution and habitat analysis of setts
4.1 Size, distribution and density of setts
Among the recorded 123 setts, the number of entrances per sett varied from one to 11 (Tab. 1). More than half the setts had only one entrance; a third of the setts had two, three or four entrances, respectively. The average number of entrances per sett was 2.26. The setts were classified into small (1 or 2 entrances), middle-sized (3-4 entrances) and big (>4 entrances) setts. Small setts were most common (88), followed by middle-sized (24) and big (11) setts.
Tab. 1: Number of entrances per sett.
|
No. of entrances |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
Total |
|
No. of setts |
72 |
16 |
10 |
14 |
1 |
3 |
0 |
1 |
4 |
0 |
2 |
123 |
|
No. of setts in % |
59 |
13 |
8 |
11 |
1 |
2 |
0 |
1 |
3 |
0 |
2 |
100 |
The distribution of the setts is shown in Fig. 8 [ISS: This figure can only be viewed with the permission of the author (Password needed)]. The average nearest neighbour distance between two setts is 111 metres, compared to the 158 metres for the average nearest neighbour distance between two generated random points. Thus, the observed distribution of the setts is significantly different from random (p<0.001). If the nearest neighbour distance is calculated for the "main setts" (setts with >2 entrances, Kruuk, 1978a) only, then the average nearest neighbour distance is 311 metres, compared to the 314 metres for the average nearest neighbour distance between two generated random points. This distribution of "main setts" does not differ from random.
The density of the setts in Sihlwald (10 km2) is 12.3/100 ha. In Fig. 9 [ISS: This figure can only be viewed with the permission of the author (Password needed)], the light coloured patches indicate areas with a higher density, dark coloured patches areas with a lower density than expected from a random distribution. Compared to the dark coloured patches, the light coloured patches "Sihlzopf", "Wüesttobel", "Schüeppenloch", "Eschentobel" and "Silrain" are very close to the forest boundary and to the adjoining agricultural areas. It is interesting to see that the setts are especially aggregated from "Häuliboden" to "Sihlzopf".
4.2 Type of sett entrances
Five different entrance types were distinguished: entrances dug directly into the ground, under a boulder/rock, under spruce trees, under deciduous trees and under tree trunks. As setts may have entrances underground as well as under spruces or under boulders, not the setts themselves, but each entrance was classified as entrance type (Tab. 2). Most entrances were dug directly into the ground ( Fig.3). The next common type was under a spruce tree ( Fig.5). It is remarkable that 31 entrances were under spruces, whereas only 7 were under deciduous trees. 22 entrances led under a boulder ( Fig.4), 12 were under tree stumps.
Tab. 2: Number of entrances for each sett type.
|
Sett type |
No. of entrances |
|
Directly into the ground |
207 |
|
Under a boulder/rock |
22 |
|
Under a spruce tree |
31 |
|
Under a deciduous tree |
7 |
|
Under a tree stump |
12 |
|
Total |
279 |
4.3 Habitat analysis of setts
Since small, middle-sized and big setts did not differ significantly from each other for any habitat parameter, all setts were taken for further analysis. The backward stepwise regression gave the best equation for the following 6 parameters: inclination, topography, distance to nearest path, middle layer-, herb- and moss coverage and explained 79.81% of the variation. Positive "B" values indicate a positive effect, negative "B" values a negative effect on sett distribution. The results show that inclination, distance to the nearest path and convex slopes have a positive effect, but flat areas, gentle and concave slopes, as well as middle layer-, herb- and moss coverage have a negative effect on sett distribution (Tab. 3).
Tab. 3: Habitat parameters that best explain the distribution of the setts. The coefficient labeled
"B" is the partial regression coefficient. Postive "B" values indicate a positive effect, negative "B" values a negative effect on sett distribution. The significant p-values (p<0.05) are printed in bold numbers. Multiple logistic regression.
|
Habitat parameter |
B |
p-value |
|
Inclination |
0.066 |
0.001 |
|
Topography |
|
0.019 |
|
flat |
-0.6172 |
|
|
gentle slope |
-0.2514 |
|
|
concave slope |
-1.5706 |
|
|
convex slope |
0.8271 |
|
|
Distance to nearest path |
0.0135 |
0.001 |
|
Middle layer coverage |
-0.0296 |
0.040 |
|
Herb coverage |
-0.3238 |
0.028 |
|
Moss coverage |
-0.708 |
0.012 |
|
Constant |
-0.1786 |
0.869 |
In Fig.9 [ISS: This figure can only be viewed with the permission of the author (Password needed)], areas with a higher sett density than expected from a random distribution are shown as light coloured patches and probably represent ideal sett sites. Therefore, a comparison was made between the setts in those areas and all other setts. However, because of the high sett concentration in the light coloured patches (Fig.9) and because of the fact that these patches are especially aggregated from "Häuliboden" to "Sihlzopf", there is a high degree of dependency between the clumped setts and the general characteristics of the southern range of the study area. Consequently, the following results should be considered with care. As the mean rank shows (Tab. 4), the setts in areas with a higher sett density than expected from a random distribution differ significantly from the other setts in the following parameters: they were found at a lower altitude (p<0.0001), had in general a lower vegetation coverage (p<0.02) and were nearer to paths than the other setts (p<0.0001).
Tab. 4: Comparison of the habitat parameters of the setts in areas with a higher density than expected from a random distribution (Bivariate Normal Kernel Density Estimation: BNKDE) with the habitat parameters of all other setts. Mann-Whitney U-test, two-tailed; mean rank, U- and p-values.
|
Habitat parameter |
|
Mean rank |
U-value |
p-value |
|
Altitude |
BNKDE setts:
other setts: |
50.36
71.73 |
1224.0 |
0.0009 |
|
Moss coverage |
BNKDE setts:
other setts: |
55.81
67.17 |
1529.5 |
0.0094 |
|
Upper layer coverage |
BNKDE setts:
other setts: |
50.15
71.90 |
1212.5 |
0.0006 |
|
Upper conifer layer coverage |
BNKDE setts:
other setts: |
53.02
69.51 |
1373.0 |
0.0087 |
|
Lower conifer layer coverage |
BNKDE setts:
other setts: |
54.90
67.93 |
1478.5 |
0.0191 |
|
Distance to nearest path |
BNKDE setts:
other setts: |
49.55
72.40 |
1179.0 |
0.0004 |
4.4 The habitat parameters
For a more detailed analysis, this chapter will compare the individual categories of the GIS habitat parameters with the availability of those categories within the study area (goodness-of-fit test). The number of setts will be compared with the number of random points for each habitat parameter measured in the field (field data) as well (Chi
2-test for two independent samples). The habitat parameters for which the distribution of the setts is non-random are then analysed with the Chi
2-test in conjunction with a Bonferroni z statistic to find out which parameter categories explain the distribution of the setts in Sihlwald best. Trends are then tested with a Mann-Whitney U-test. It should be kept in mind that, with this number of parameters, the probability of a random significance is high. Fig. 10-21 help to clarify the ecological significance of the parameters. Therefore, the significance of the individual classes is less relevant than the pattern of each parameter. Likewise, the individual parameters are only parts of a more complex system of habitat factors influencing the choice of sett sites in Sihlwald.
The distribution of the setts is decidedly non-random for the following GIS parameters (Tab. 5): altitude (p<0.001) and distance to the nearest path (p<0.001) seem to have the biggest effect on the distribution of the setts. The vegetation coverage in general seems to play a key role. The distribution of the setts also depends on aspect (p<0.02) and distance to nearest water (p<0.05). The parameter categories affecting the distribution of the settts are shown below (Fig. 10-16). No significant difference was attained for the following parameters: upper- and middle layer coverage, upper coniferous layer coverage, vegetation unit and development stage of the forest stands.
Tab. 5: Comparison of the GIS habitat parameters of the setts with the availability of those parameters within the study area. Only the parameters for which the distribution of the setts is significantly non-random are listed here. Goodness-of-fit test; df= degrees of freedom.
|
Habitat parameter |
p-value |
Chi
2 |
df |
|
Altitude |
p<0.001 |
5046.96 |
3 |
|
Aspect |
p<0.02 |
16.78 |
7 |
|
Lower layer coverage |
p<0.02 |
12.69 |
4 |
|
Lower coniferous layer coverage |
p<0.005 |
12.39 |
2 |
|
Middle coniferous layer coverage |
p<0.02 |
10.89 |
3 |
|
Distance to nearest path |
p<0.001 |
18.10 |
3 |
|
Distance to nearest water |
p<0.05 |
10.82 |
4 |
Setts are found significantly closer to the forest boundary than random points (Mann-Whitney U-test, two tailed; U=4003, p<0.005). Comparison of the habitat parameters of the setts measured in the field with those of the random points (Tab. 6) shows that setts differ significantly from random points in the following parameters: inclination, topography (both p<0.001), herb- (p<0.05) and moss coverage (p<0.01). No significant difference was attained for shrub coverage. The parameter categories affecting the distribution of the setts are shown below (Fig. 17-21). Categories containing less than 3 data points were pooled.
Tab. 6: Comparison of the habitat parameters of the setts with those of the random points. Only the parameters for which the distribution of the setts is significantly non-random are listed here.
chi2-test for two independent samples; df=degrees of freedom.
|
Habitat parameter |
p-value |
Chi
2 |
df |
|
Inclination |
p < 0.001 |
52.41 |
4 |
|
Topography |
p < 0.001 |
40.04 |
4 |
|
Herb coverage |
p < 0.05 |
19.63 |
5 |
|
Moss coverage |
p < 0.01 |
15.44 |
2 |
As shown in Fig. 10, the number of observed setts differs significantly from the number of expected setts (p<0.05) for each altitude category. There are fewer setts than expected below 600 m a.s.l., but more than expected above 600 m a.s.l. (Mann-Whitney U-test, two tailed; U=0, p<0.001).
The observed and expected number of setts do not differ for any aspect category other than the category North, for which fewer setts than expected were found (p<0.05; Fig. 11).
No trend can be seen for lower layer coverage (Fig. 12). Setts are preferably dug in areas lacking a lower- and middle coniferous layer coverage (Fig. 13, Fig. 14), rather than in areas containing a lower- and middle layer coverage (both: Mann-Whitney U-test, two tailed; U=0, p<0.001).
As shown in Fig. 15, there were significantly fewer setts than expected in category £50 m (p<0.05), but more than expected in the category 51-100 m to the nearest path (p<0.05). In general, fewer setts are found £50 m to nearest path than >50 m to the nearest path (Mann-Whitney U-test, two tailed; U=0, p<0.001). No trend can be seen for any category of distance to nearest water (Fig. 16). Significantly more setts than expected were found in the distance category 51-100 m (p<0.05) from the forest boundary (Fig. 17), fewer than expected in the distance categories 301-400 m and >900 m from the forest boundary (p<0.05).The difference between setts found <=
300 m and those found >300 m from the forest boundary is significant (Mann-Whitney U-test, two tailed; U=0, p<0.001).
Setts are rarely dug in flat areas (p<0.05; Fig. 18). They are preferably dug on slopes with an inclination <=
20°, rather than on slopes with an inclination >20° (Mann-Whitney U-test, two tailed; U=38.5, p<0.001). Topography plays a key role in the choice of sett site (Fig. 19): convex slopes are the preferred sett locations (p<0.05) whereas flat areas, gentle and concave slopes are avoided (p<0.05).
As shown in Fig. 20, setts are preferably dug in areas with a sparse herb coverage, rather than in areas with a dense herb coverage (Mann Whitney U-test, two tailed; U=0, p<0.001). The same trend can be seen in Fig. 21: areas without moss are significantly preferred to areas with moss (Mann-Whitney U-test, two tailed; U=0, p<0.001). See Appendix , (Tab.18-29) for exact data on Chi
2-test in conjunction with a Bonferroni z statistic.
4.5 Discussion of the sett distribution, density and types of entrances
The recorded setts vary greatly in size, extent of tunnels and the number of entrances. Possible explanations for these observations are:
• setts of different sizes are used for different purposes (Thornton, 1988);
• setts are larger and have more entrance in regions where the soil is more easily dug (Neal, 1977);
• the size of a sett, especially of a main sett, reflects its age, since badgers tend to go on extending their setts for as long as the occupation continues (Neal & Roper, 1991).
Since neither size of the setts nor extent of the tunnels were examined in this study, the setts in Sihlwald were classified in small, middle-sized and big setts according to the number of entrances. The number of entrances per sett varied from one to 11 with an average of 2.26 entrances per sett. This is well below the average found in the literature: Kruuk (1978a) found main setts with three to 21 entrances and outliers with one or two entrances resulting in an average of 10.5 entrances per sett. In Belgium, the number of setts varied from one to 38 entrances with an average of 13.3 entrances per sett (Anrys & Libois, 1988). This difference can be explained by the fact that this study did not differentiate between fox dens and badger setts. Fox dens usually have fewer entrances than badger setts (Stubbe, 1980). This could be the reason why the average numbers of sett entrances in Sihlwald is lower than the average mentioned by other authors. Nevertheless, badger hairs were found in several setts consisting of one entrance only, indicating the presence of badgers in single entrance setts as well. As mentioned before, it was seldom possible to clearly distinguish between setts used by badgers and those used by foxes. The differentiation was especially difficult for setts with 1 or 2 entrances. Badger hairs indicated that a badger had visited the sett, but gave no indication for how long or how often the particular sett was used. Tracks were only found on freshly dug spoil heaps or around the entrance after it had rained. So, owing to this uncertainty concerning most of the setts, all setts (unless mentioned elsewhere) were used for further analysis.
The setts in Sihlwald were non-randomly spaced. The average nearest neighbour distance was 111 metres. Since too little is known about the utilisation of the different setts, they could not be classifed into main setts and outliers. Nevertheless, in order to compare the nearest neighbour distance between the main setts as defined in Kruuk (1978a), all setts with more than 2 entrances were taken for further analysis. The distribution of the main setts did not differ from random and the average nearest neighbour distance between two main setts was 311 metres. The average nearest neighbour distance corresponds fairly well with that of the main setts in southern England (300 metres, Kruuk, 1978a), although the distribution of those setts was non-random.
The density of setts in Sihlwald is 12.3/100 ha. Clements et al. (1988) think that 10 setts/100 ha is an average sett density. Compared to the low densities in former Czechoslovakia (0.18/100 ha, Pelikan & Vackar, 1978) and in Switzerland (Canton of Neuchatel: 0-0.22/100 ha, Monnier, 1993; Canton of Berne: 0.42/100ha, Graf, 1988) and the relatively high sett densities all over Great Britain (0-26/100 ha, Cresswell et al., 1990), Sihlwald seems to have an average sett density for European standards. However, for Swiss standards Sihlwald has a high sett density.
The areas "Sihlzopf", "Wuesttobel", "Eschentobel", "Schüeppenloch" and "Silrain" have a higher sett density than expected from a random distribution. These areas are also very close to the forest boundary and to the adjoining pastures and agricultural zones (see also 4.6). In the Canton of Zurich, badger setts are mainly found in the richly structured forest- and agricultural zones (Hindenlang, 1992). This mixture of woodland and pastures, woodland and arable land is among the habitat types preferred by the badger (Neal, 1977; Zejda & Nesvadbova, 1983). Blab et al. (1989) mention two possible reasons: 1. the topography in such regions is extremely varied, which is advantageous for building setts on slopes; 2. important food resources (earthworms, insects) are found in higher densities in this diverse landscape. Neal (1986) also thinks that the presence of a plentiful and varied food supply is one of the most important factors influencing the choice of a sett site. If this is true, the areas in Sihlwald with a higher sett density than expected from a random distribution are ideal sett sites. The setts are situated close to pastures and arable land, meaning that badgers can adapt their foraging strategies to seasonal changes in food availability in the different habitats.
The area "Häuliboden" is an exception. It is not close to any forest boundary, yet there are several setts in this area. It seems that the biggest sett there has been used for generations and is an ideal sett site regarding the heterogeneous topography and sparse vegetation cover.
Areas with a lower sett density than expected from a random distribution (dark coloured patches, Fig. 9) are relatively flat, are close to the village "Langnau a. A." and are therefore near frequently used paths. There is also a higher hunting pressure in this region than toward Sihlzopf. Thus, these areas are less attractive sett sites.
In this study, five different entrance types were
distinguished according to the location of the entrances: entrances
dug directly into the ground, under a boulder/rock, under a spruce
tree, under a deciduous tree and under a tree stump. Although setts were analysed in several studies, only Bock (1986) classified setts in a similar way. Most of the setts were also dug straight into the ground. Bock mentions however, that in montane regions, most setts built underground are actually under a rock. In Sihlwald, 50 entrances were built under trees (tree stumps were counted here as trees, since it is not clear whether the trees were cut down before or after the entrances were dug). It is surprising that 31 entrances were dug under spruces. This has never been mentioned in the literature before. These spruces were all in mixed forest stands (with two exceptions in coniferous stands; both were fox dens). A possible explanation is that spruce trees are normally shallow rooted (Köstler et al., 1968; Blanckmeister & Hengst, 1971), compared to the dominant beech trees in Sihlwald. This facilitates digging and has the advantage that the roots keep the roof of the sett from caving in. For forestry management considerations, this means that single spruces should be left standing in the individual forest stands.
4.6 Discussion and synthesis of the habitat characteristics of the setts
This study was centered on finding habitat
parameters likely to be relevant to the distribution of sett and
latrine sites. Yet, it is often impossible to separate the relative
importance of various ecological parameters,as they are obviously
inter-related and differ in priority from one area to another. This
study probably does not contain all relevant parameters for a sett
site for two reasons: 1. it is difficult to know beforehand which
parameters are likely to be relevant; and 2. the more parameters are
measured, the more difficult it is to find strong relationships
between parameters and the more difficult it gets to interpret the
results correctly.
The first parameter to be discussed here is the type of soil. This parameter seems to be important in the choice of a sett site (Neal, 1986; Dunwell & Killingley, 1969; Zejda & Nesvadbova, 1983). No exact data was collected on different soil types at each sett. Information about soil came from the corresponding map (Bodeneignungskarte des Kantons Zürich, 1: 50'000). The dominant soil types in Sihlwald are sandy to silty clay and argillaceous sand. Sandy soil is normally preferred to one of clay (Neal, 1986). The only disadvantage of sandy soils is that if they are very light there is a risk of the roof of the sett collapsing. Neal (1986) mentions, however, that these soils are often associated with woodland and that badgers choose sett sites where tree roots provide the necessary strength. In Sihlwald, setts built under tree roots were seen on several occasions (Fig. 4) (see also 5.3). All in all, Sihlwald seems to have an ideal soil for digging setts (see also topography, p. 22). This factor could possibly explain the relatively high sett density in this forest compared to other densities in Switzerland (see above).
Altitude: fewer setts than expected were found below 600 metres a.s.l., more setts than expected were found above 600 metres. A possible reason for this observation could be the proximity of the "Sihltalstrasse" in the lower ranges, the main road connecting Zurich with Central Switzerland. This road is permanently busy and represents along with the railroad and the river Sihl, a major barrier for the animals. It is unclear whether it is the noise of the passing cars and trucks or the level of traffic mortality that keeps the badgers from digging their setts in proximity of the road (Skinner et al., 1991). However, only one or two badgers are killed on the "Sihltalstrasse" yearly (Leutwyler, pers. comm.) As already mentioned above, it is likely that food distribution (see also distance to forest boundary, p. 23) plays a bigger role in the distribution of the setts than the noise of passing vehicles or traffic mortality.
Aspect: significantly fewer setts than expected faced North. However, no preference was attained for any other category. In general, slopes facing north seem to be unattractive sett sites (Monnier, 1993; Skinner et al., 1991; Anrys & Libois, 1983). Owing to the orientation of the forest (SSE to NNW) and thus to lack of slopes facing south, a preference for this aspect could not be confirmed during the study.
Inclination: most setts in Sihlwald were dug in a slope with an inclination between 20°-40°. Fewer than the number of expected setts were dug in flat areas (£10°). Setts are usually dug in slopes of some sort (Neal, 1986; Skinner et al, 1991). Thornton (1988) showed that main sett density increased with the hilliness of the district. This is advantageous to the badger in various ways (Neal, 1986):
• digging into a slope facilitates the removal of the excavated soil, which spills down the slope;
• it is easier for the badger to find a particularly favourable stratum of soil for digging since it is more likely to be exposed;
• sloping land is usually well drained so that the sett is more likely to be warm and dry;
• and in colder parts, a depth below ground is quickly attained which is frost proof.
Topography: this parameter is closely connected with inclination. Setts in Sihlwald were preferably dug in convex slopes. Such setts have the following advantages: the badgers can pick up scents from several directions without having to leave the security of the sett and thus have several directions from which they can leave the sett. It is also possible that setts on convex slopes are easier to enlarge, since the rounded shape of the slope gives the badgers more opportunities for digging entrances and connecting tunnels than the shape of a gentle slope. Topography is a parameter that has never been mentioned in the literature before and seems to play a key role in the choice of a sett site. It appears to be important to pay attention to the small-scale topography around the setts.
Vegetation cover in general: the preferred sett sites are tree stands with sparse ground cover (i.e. a low herb- and moss coverage). As mentioned in Neal (1986) and Zejda & Nesvadbova (1983), coniferous stands were avoided. In Sihlwald, badgers clearly preferred areas with a sparse lower- and middle coniferous layer coverage. However, as shown above (4.5), single spruces were popular sett sites. According to Neal (1986), cover seems to play an important role in the choice of a sett site. Cover allows the badgers to emerge inconspicuously, and it allows the young cubs to play near the entrance without being visible to people or potential predators. However, the parameter cover is never clearly defined. Cover does not only have to be the vegetation cover; the small-scaled topography around a sett can be a type of cover, too. The distance to the nearest path (just "being out of view") can indirectly be a type of cover as well. It should be mentioned that no comparisons were made between the field data tree coverage and the lower-, middle-, and upper layer coverage, since the data of the latter are averages of the individual forest stands and thus too vague to be compared with the exact data of tree coverage at each site.
Distance to nearest path: Sihlwald is a popular recreation area for joggers, mountain bikers and hikers, especially on weekends. Fortunately, owing to the topography and the often dense vegetation of this forest, people rarely leave the paths. However, even though badgers are nocturnal, they avoid digging their setts close to the paths. Setts are most common in the distance categories >50 metres to the nearest path.
Distance to nearest water: there is no area in Sihlwald that is very far from water. Thus, it is assumed that this is the reason why no significant difference was attained for any distance category to nearest water. It is not clear why this parameter affects the distribution of the setts, but it probably correlates with other habitat parameters relevant to sett sites.
Distance to forest boundary with adjoining agricultural zones: setts are found significantly closer to the forest boundary than random points. Ideal sett sites are determined by at least two factors: the proximity to the forest boundary and adjoining agricultural zones, guaranteeing the presence of a good food supply on the one hand, and the forest itself, providing slopes, cover and little disturbance on the other hand.
The setts found in areas with a higher sett density than expected from a random distribution (light coloured patches, Fig. 9 were generally situated at a lower altitude, had a lower vegetation coverage and were nearer to paths than the other setts. This can be explained by the fact that the area from "Häuliboden" to "Sihlzopf" is actually at a lower altitude than the rest of the forest. The forest is narrower in this area, so that the setts were closer to the paths and to the forest boundary.
In order to understand the results of the analysis of the setts mentioned above, Fig. 22 shows the different parameters affecting sett sites in Sihlwald.
5 Distribution and habitat analysis of latrines
5.1 Size, utilisation, distribution and density of latrines
During the survey 40 latrines were recorded. The number of dung pits per latrine varied from one to 20 (Tab. 7). Nearly half the latrines only had a single pit. Four latrines had three dung pits, two had four, four had five, and two had six dung pits. The biggest latrine consisted of 20 dung pits. The average of dung pits per latrine was 3.5. The number of defecations varied from one to seven, with an average of 1.475 defecations per latrine.
Tab. 7: Latrine size and utilisation.
|
No. of latrines |
19 |
1 |
4 |
2 |
4 |
2 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
|
No. of dung pits |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
11 |
12 |
14 |
16 |
19 |
20 |
Sum
: 140; Avg
: 3.5 |
|
No. of used dung pits |
1 |
2 |
1-3 |
3-4 |
3-5 |
3 |
5 |
7 |
3 |
4 |
4 |
2 |
7 |
2 |
Sum
: 50; Avg
: 1.475 |
Fig. 23 [ISS: This figure can only be viewed with the permission of the author (Password needed)] shows the distribution of the latrines in Sihlwald. Since only 40 latrines were found, the distribution of the latrines was not tested against a random distribution. All that can be said is that the minimum density of latrines in Sihlwald is 4/100 ha.
5.2 Latrine location
There is a significant difference between the latrines near (<=
5 m) setts and those that are not in the immediate vicinity of a sett. As the mean rank shows (Tab. 8), latrines near setts contain less dung pits (p=0.0103) and less used dung pits (p=0.0377) than latrines further away from setts.
Tab. 8: Comparison of the number of dung pits and used dung pits of latrines near setts with those not near setts. Mann-Whitney U-test, two-tailed; mean rank, U- and p-values.
|
Latrine composition |
|
|
Mean rank |
U-value |
p-value |
|
Dung pits |
Avg
: 1.67
Avg
: 5.71 |
near setts:
not near setts: |
13.67
23.43 |
86.0 |
0.0103 |
|
Used dung pits |
Avg
: 1.58
Avg
: 2.75 |
near setts:
not near setts: |
15.04
22.84 |
102.5 |
0.0377 |
Too few latrines were found to test the following details. Nevertheless, certain observations are worth noting: of the 40 latrines found, 23 had a clear runway nearby. More than two thirds (28) of the latrines were found near (<=
5 m) a conspicuous landmark (Tab. 9). The most commonly used landmark was near a sett, followed by near a path. Four latrines were found at the edge of the forest (one of which was near a fence post), two on the border of two different forest stands (mixed=deciduous and coniferous trees/deciduous trees) and two on the border of two development stages (old mixed/young deciduous stand). One latrine was found in an entrance of a sett. A third of the latrines (12) could not be placed in any of the above categories.
Tab. 9: Number of latrines near different types of landmarks.
|
Type of landmark |
No. of latrines |
|
Near a sett |
11 |
|
In a sett |
1 |
|
Near a path |
8 |
|
Forest boundary |
4 |
|
Border of two different stands |
2 |
|
Border of two development stages |
2 |
|
No conspicuous landmark |
12 |
|
Total |
40 |
5.3 Habitat analysis of latrines
The GIS habitat parameters of the latrines for which the distribution of the latrines is decidedly non-random are listed in Tab. 10. Altitude seems to have the biggest effect on the distribution of the latrines (p<0.001), followed by distance to nearest water (p<0.05).
No significance was attained for any habitat parameter in the Chi2-test for two independent samples (latrines versus random points).
Significantly fewer latrines than expected were found below 600 m a.s.l. (p<0.05) (Fig. 24).
The difference between the number of observed and the number of expected setts was significant for the category >800 m a.s.l. (p<0.05). Latrines were more often found in areas >600 m a.s.l., than in areas £ 600 m a.s.l. (Mann-Whitney U-test, two tailed; U=0, p<0.001). No significant difference was attained for any distance category to nearest water (Fig. 25). Latrines were found significantly closer to the forest boundary than random points (Mann-Whitney U-test, two tailed; U=1316.5, p<0.05). See Appendix (Tab. 30 & 31) for exact data on Chi2-test in conjunction with a Bonferroni z statistic.
Tab. 10: Comparison of the GIS habitat parameter categories of the latrines with the availability of those parameter categories within the study area.
|
Habitat parameter |
p-value |
chi 2 |
df |
|
Altitude |
p<0.001 |
6132.53 |
3 |
|
Distance to nearest water |
p<0.05 |
10.82 |
4 |
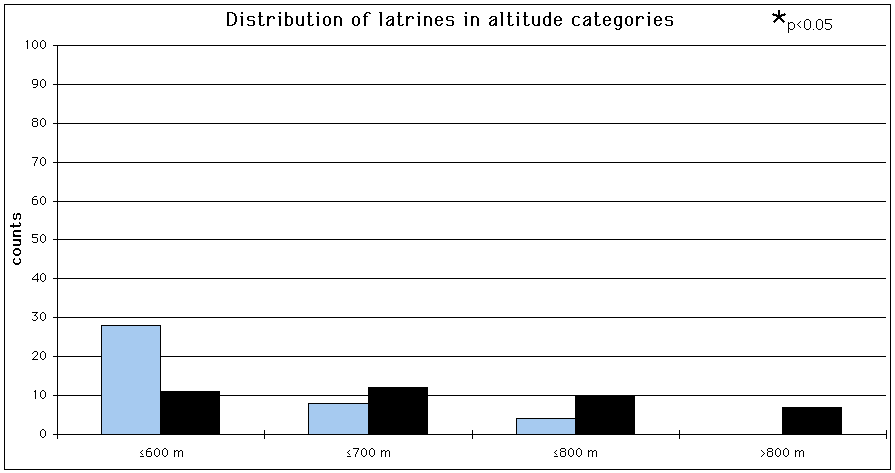
Fig. 24: Distribution of the latrines for altitude categories (Bonferroni z statistic, Neu et al., 1974). Dark coloured bars = number of setts, light coloured bars = number of random points.
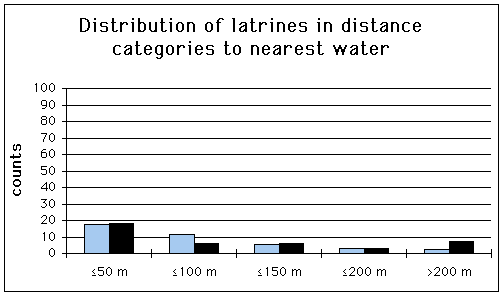
Fig. 25: Distribution of the latrines for categories to nearest water. (Bonferroni z statistic, Neu et al., 1974). Dark coloured bars = number of setts, light coloured bars = number of random points.
5.4 Discussion of the distribution, density and utilisation of latrines
Within 40 latrines recorded, the number of dung pits per latrine varied between one and 20, with a mean size of 3.5 pits per latrine. The number of defecations per latrine varied from one to seven, with an average of 1.475 defecations per latrine. Roper et al. (1991) mention shared latrines (boundary latrines) with one to 15 dung pits and a mean size of 4.4 pits per latrine. Unshared latrines ( boundary latrines) had one to three pits and a mean size of 2.6 pits per latrine. Shared latrines contained an average of 8.7 defecations per latrine, unshared latrines only an average of 4.2 deposits. Taking the fact that the two types of latrines were not distinguished in Sihlwald, my results regarding mean size of dung pits correspond fairly well. However, the average defecations per latrine is much lower than the average mentioned in Roper et al. (1991). This can be explained by the fact that every latrine in Sihlwald was only recorded once, meaning that these results reflect a chance sample in utilisation.
The density of latrines in Sihlwald is at least 4/100ha. It is certain that many latrines were overlooked. Comparisons with Kruuk (1978a): 31/100ha and Roper et al. (1991): 37/100ha show that the density of the latrines is probably higher regardless of possible differences in animal density and spatial organisation. Furthermore, latrine use seems to be seasonal (Kruuk, 1978a; Roper et al., 1991; Pigozzi, 1990) with peaks in spring (Feb.-May) and to a lesser extent in autumn (Sept.-Oct.). It is possible that only few latrines were found due to the time of mapping in Sihlwald (May-Aug.). Nevertheless, looking back at the habitat characteristics (topography, inclination, vegetation cover) of
the study area, it is still unclear which method would have revealed more latrines.
Latrines near setts had significantly fewer dung pits and fewer used dung pits than latrines elsewhere. Kruuk (1978a) found that latrines on territorial boundaries were larger than those near setts. Latrines on boundaries have a territorial function, whereas latrines elsewhere are used for defecation only (Roper et al., 1986). This might signify that some latrines in Sihlwald have a territorial function. It is also assumed that, compared to the wide seasonal variation in food availability in the agricultural zones, the forest has a more predictable food availability throughout the year. This would again point to a territorial social system. However, Packham (1983) found that badgers in the forest made no attempt to delimit range boundaries and that latrines appeared to be distributed randomly within the animals’ range. This is possibly due to the low earthworm density within the forest, which results in prediction of a range so large it would be impossible to defend. Thus, according to Packham (1983), badgers with home ranges exclusively in the forest should be more or less solitary, with extensive, overlapping home ranges. It should be kept in mind however, that earthworm densities depend of the type of forest. Deciduous forest stands can have the same earthworm densities as certain pastures (Thomas Nabulon, pers. comm.). It remains to be seen whether or not latrines have a territorial function in Sihlwald.
Of the 40 latrines found in Sihlwald, 28 were near some kind of conspicuous landmark (Tab. 10). Of the 164 latrines mentioned in Kruuk (1978a), 129 were near a landmark. However, these results cannot be directly compared since different types of landmarks were defined. Nevertheless, latrine sites in Sihlwald seem to be near certain conspicuous landmarks as well. Landmarks are usually used when places in an animal’s range are returned to time and time again. This would be the case if latrines had a territorial function. More information is needed about the badgers spatial and social organisation before further analysis can be made with regard to different latrine types and their correlation to landmarks.
5.5 Discussion of the habitat analysis of latrines
The distribution of the latrines in Sihlwald seems to be determined by altitude and distance to nearest water. There appears to be a correlation between altitude and distance to agricultural zones, meaning that altitude may not be the parameter actually determining the distribution of the latrines. The latrines are found in areas where there is a special abundance of food (Neal & Cheeseman, 1996). This would explain why there are significantly fewer latrines than expected below 600 metres a.s.l. (near the main road) and significantly more than expected above 600 metres a.s.l. (near to pastures and agricultural zones). This is further confirmed by the fact that latrines are significantly closer to the forest boundary than random points. Some badgers go out of the forest to feed in the pastures (Stefan Imfeld, pers. comm.) and may stop at latrines on their way back to the sett. As already mentioned for the setts, there is no area in Sihlwald that is very far from water. For this reason no significant difference was attained for any distance category to nearest water. It is assumed that this parameter correlates with other habitat parameters relevant to sett sites.
Sett and latrine sites are related, the later being in some way determined by the former. Thus, it is not surprising to see that some parameters responsible for the distribution of the setts are also responsible for the distribution of the latrines: altitude and distance to forest boundary, both correlating in some way with distance to agricultural zones, and distance to nearest water, seem to play a key role in the spatial organisation of the badger.
This habitat analysis should be considered as a trial. Too few latrines were found to obtain any significant results. More information is needed about the badger's habitat utilisation in general, before further studies on latrines are carried out.
6 Sihlwald - an ideal badger habitat
The high sett density in this forest indicates an ideal badger habitat. This forest has a pronounced topographical variation: the convex slopes are ideal sett sites. They are well drained and offer many opportunities for digging entrances and tunnels, and thus offer several directions from which a badger can leave the sett. Sihlwald is characterised by a diverse pattern of mixed forest, dominated by deciduous trees. The single spruces in these forest stands are important sett sites. Their shallow roots facilitate digging and help keep the roof of the setts from caving in. The different forest stands guarantee a good food supply all year round; the adjoining agricultural zones offer a wide seasonal variation in food availability.
The size of the setts in Sihlwald may indicate differences in sett use: a radio-collared female badger is known to use a certain big sett regularly, but visits other small ones from time to time (Karin Hindenlang, pers. comm). The distribution of setts and latrines indicates that certain badgers probably leave the forest to forage for food, whereas data of the radio-collared badgers show that certain individuals never leave the forest (K. Hindenlang, pers. comm.). This leads to the assumption that several strategies may exist side by side: from individuals that live in groups, have distinct home ranges and occupy main setts, to individuals that are solitary, have large overlapping home ranges, forage outside the forest and occupy several setts within a range. It remains to be seen, just how flexible the social and spatial organisation of the badgers in Sihlwald is.
7 References
Anrys P. & R. M. Libois (1983): Choix de l'habitat chez le blaireau européen (Meles meles) en Bélgique. Cahiers d'Ethologie apliquée 3 (1): 15-38.
Bgu (1988): Naturlandschaft Sihlwald, Studienbereich A, Vegetation.
Blab J., A. Terhardt & K.P. Zsivanovits (1989): Tierwelt in der Zivilisationslandschaft. Schriftreihe für Landschaftspflege und Naturschutz, Heft 30. Bonn-Bad Godesberg.
Blankmeister J. & E. Hengst (1971): Die Fichte im Mittelgebirge. Neumann Verlag.
Bock W. F. (1986): Kriterien zur Grössenbeurteilung der Baue des Dachses (Meles meles L.). Säugetierkundliche Mitteilungen 33: 227-234.
Bodeneignungskarte der Schweiz (1980): Blatt 225: 1:50'000. Bundesamt für Raumplanung.
Braun-blanquet J. (1964): Pflanzensoziologie. Springer Verlag, Wien, New York.
Bundesamt für Landestopographie, 3084 Wabern (1988): Landeskarte der Schweiz. ALBIS 1111. 1:25'000.
Cheeseman C. L., W. J. Cresswell, S. Harris, & P. J.Mallinson (1988): Comparison of dispersal and other movements in two badger (Meles mels) populations. Mammal Rev. 18 (1):51-59.
Clements E. D., E. Neal & D. W. Yalden (1988): The National Badger Sett Survey. Mammal Rev., Vol. 18, No. 1: 1-9.
Cresswell W. J. & S. Harris (1988): Foraging behaviour and home range utilisation in a suburban badger (Meles meles) population. Mammal Rev. 18 (1): 37-49.
Cresswell P., S. Harris & D. J. Jefferies (1990): The history, distribution, status and habitat requirements of the badger in Britain. Nature Conservancy Council, Peterborough.
Dunwell M. R. & A. Killingley (1969): The distribution of badger setts in relation to the geology of the Chilterns. J. Zool. 158: 204-208.
Graf M. (1988): Die räumliche und zeitliche Habitatnutzungeiner Dachspopulation am Gurten bei Bern. Dissertation Univ. Bern.
Graf M. (1995): Meles meles in J. Hausser (ed.): Säugetiere der Schweiz (1995): Birkhäuser Verlag, Basel, Boston, Berlin.
Hantke R. (1967): Geologische Karte des Kanton Zürichs und seiner Nachbargebiete. In zwei Blättern 1: 50'000. Kommissionsverlag Leemann, Zürich.
Hindenlang K. (1992): Analyse und Beurteilung der Landschaft im Kanton Zürich anhand der Verbreitung ausgewählter einheimischer Wildtierarten. Diplomarbeit Univ. Zürich.
Imfeld S. (in prep.): How to catch running animals with a GIS....and see what they’re up to.
Köstler J. N., E. Brückner & H. Bibelriether (1968): Die Wurzeln der Waldbäume. Untersuchungen zur Morphologie der Waldbäume in Mitteleuropa. Verlag Paul Parey. Hamburg und Berlin.
Kruuk H. (1978a): Spatial organisation and territorial behaviour of the European badger, Meles meles. J. Zool. Lond. 184: 1-19.
Kruuk H. (1989): The social badger. Oxford University Press.
Monnier M.-F. (1993): Le blaireau (Meles meles L.) dans le canton de Neuchâtel (Suisse). Travail de diplôme. Univ. Neuchâtel.
Mosteller F. & J.W. Tukey (1977): Data analysis and regression. A second course in statistics. Addison-Wesley Publishing Company.
Neal E. (1977): Badgers. Blandford Press Ltd., Poole, Dorset (first edition: 1949).
Neal E. (1986): The natural history of badgers. Croom Helm, London.
Neal E. & T. J. Roper (1991): The environmental impact of badgers (Meles meles) and their setts. Symp. Zool. Soc. Lond. No. 63: 89-106.
Neal E. & C. Cheeseman (1996): Badgers. T & AD Poyser Ltd. Natural History.
Neu C.W. & C.R. Byers & J.M. Peek (1974): A technique for analysis of utilisation-availability data. J.Wildl.Manage. 38(3): 541-545.
Packham C. G. (1983): The influence of food supply on the ecology of the badger. BSc. Honours thesis (Biology), University of Southhampton.
Pelikan J. & J. Vackar (1978): Densities and fluctuation in numbers of red fox, badger and pine marten in the "Bucin" forest. Folia Zool. 27 (2): 289-303.
Pigozzi G. (1990): Latrine use and the function of territoriality in the European badger, Meles meles, in a Mediterranean coastal habitat. Anim. Behav. 39(5): 1000-1002.
Roper T. J. (1992): Badger Meles meles setts-architecture, internal environment and function. Mammal Rev.,Vol. 22, No. 1: 43-53.
Roper T. J. (1992): The structure and function of badger setts. J. Zool. Lond. 227 (4): 691-694.
Roper T. J., A. I. Tait, D. Fee & S. F. Christian (1991): Internal structures and contents of four badger (Meles meles) setts. J. Zool. Lond. 225: 115-124.
Roper T. J. , D. J. Shepherdson & J. M. Davies (1986): Scent marking with faeces and anal secretion in the European badger (Meles meles): seasonal and spatial characteristics of latrine use in relation to territoriality. Behaviour 97: 94-117.
Sachs L. (1984): Nichtparametrische statistische Methoden. Frankfurt: Fachbuchhandlung für Psychologie, Verlagsabteilung.
Skinner C. A. & P. J. Skinner (1988): Food of badgers (Meles meles) in an arable area of Essex. J. Zool. Lond. 215: 360-362.
Skinner C., P. Skinner & S. Harris (1991): An analysis of some of the factors affecting the current distribution of badger (Meles meles) setts in Essex. Mammal Rev., Vol. 21, No. 2: 51-65.
Stadtforstamt Zürich (1990): Bestandeserhebung 1990.
Stubbe M. (1980) Population ecology of the red fox (Vulpes vulpes L.) in the GDR. Biogeographica Vol. 18: The Red Fox.
Thornton P. S. (1988): Density and distribution of badgers in south-west England - a predictive model. Mammal Rev., Vol. 18, No. 1: 11-23.
Weber D. (1985): Zur Baubenutzung und ihrer Funktion beim Fuchs (Vulpes vulpes L.). Z. Säugetierkunde 50: 356-368.
Weber D. (1990): Aspekte der Nutzung von Unterschlüpfen beim Fuchs. Symp. AKW. 20: 23-25.
Zejda J. & J. Nesvadbova (1983): Habitat selection and population density of the badger (Meles meles) in Bohemia and Moravia. Folia Zool. 32: 319-333.
{kind=link}
{kind=link}
{kind=link}