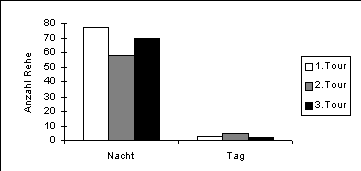
In der Nacht wurden auf der gesamten in Abbildung 2 gezeigten Beobachtungsfläche von ca. 2 km2 im Durchschnitt 68.3 Rehe pro Durchgang gesehen. Im Vergleich dazu wurden am Tag jeweils nur rund 3 Rehe beobachtet (Abbildung 5). Dieser Unterschied veranschaulicht eine deutliche Ausweichreaktion der Rehe am Tag weg von den grösseren Wegen. Trotz der kleinen Stichprobe am Tag zeigt sich eine Tendenz, dass Rehgruppen am Tag in steileren Lagen beobachtet wurden (MWU, n1=116, n2=7, z=-1.74, p=0.08). Dagegen lässt sich kein Unterschied in der Höhenverteilung nachweisen (MWU, n1=116, n2=7, z=-0.699, p=0.48).
Um zu prüfen, ob zwischen den drei Beobachtungsaufnahmen in der Nacht Unterschiede auftraten, wurden die Parameter in Tabelle 4 mittels H-Tests auf Unterschiede in ihrer zentralen Tendenz geprüft. Dabei zeigten sich weder bei der Höhenverteilung, den Distanzen zu den Futterkrippen und Gewässern signifikante Unterschiede.
| Parameter | Chi2 | p |
| Höhe | 1.36 | 0.51 |
| Distanz zur Futterkrippe | 0.20 | 0.90 |
| Distanz zu Gewässern | 0.46 | 0.79 |
| Hangneigung | 5.05 | 0.08 |
| Gruppengrösse | 5.39 | 0.07 |
Bei der Hangneigung zeigte sich eine Tendenz zu steileren Gebieten im Übergang von der ersten (Durchschnitt 9.3) zur zweiten Aufnahme (Durchschnitt 12.0). Ebenfalls nur als Tendenz erwies sich eine Abnahme der Gruppengrösse. Die durchschnittliche Anzahl Rehe pro Gruppe sank von 2.0 in der ersten auf 1.5 Tiere in der dritten Aufnahme, was darauf zurückzuführen ist, dass vermehrt Einzeltiere und weniger Zweiergruppen zu beobachten waren (Abbildung 6).
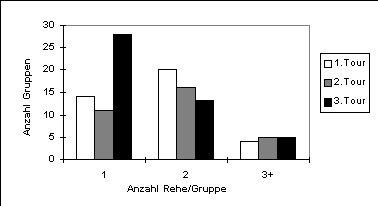
Die Anzahl Rehe, die auf den Tagestouren gesehen wurden, ist zu gering, um eine vernünftige Analyse der Einflussfaktoren durchzuführen. Deshalb beschränke ich mich im folgenden auf die Nachtbeobachtungen. Dies auch in der Annahme, dass die Verteilung der Rehe in der Nacht sehr viel weniger durch die menschlichen Aktivitäten beeinflusst wird. Die Beobachtungsorte der Rehe während der Nacht sind in Abbildung 7 dargestellt. Sie sind nicht gleichmässig über das ganze Gebiet verteilt, sondern bilden grob drei Zentren in den Gebieten (1) Schönenboden, (2) Biri-, Schröter- und Häuliboden und (3) Cholbenholz (vgl. Abbildung 2). Zum anderen fällt eine grosse Lücke ohne Rehbeobachtungen auf, die ungefähr durch das Dreieck Spinnerweg-Waldmattstrasse/Weierbrunnenstrasse-Albishornstrasse eingefasst wird. Im folgenden gehe ich auf einzelne Faktoren ein, die die Verteilung beeinflussen könnten, und danach auf die multivariaten Verfahren für die Extrapolation der Verteilung auf den gesamten Sihlwald.
A: Chi2-Anpassungstests mit simultanen Konfidenzintervallen (Bonferoni z-Statistik)
Die Ergebnisse der Chi2-Anpassungstests für die einzelnen Parameter sind in Tabelle 5 zusammengestellt. Die Verteilung der Rehgruppen bezüglich Hangneigung unterscheidet sich signifikant vom Angebot. Die Hangneigungsklassen über 20, die rund 24% der Beobachtungsfläche ausmachen, weisen signifikant weniger Beobachtungen als erwartet auf. Bei der Höhenverteilung, die sich ebenfalls signifikant vom Angebot unterscheidet, weist die höchstgelegene Klasse der Beobachtungsfläche von 750-830m signifikant weniger Rehgruppen auf. Die Klasse 630-670m weist signifikant mehr Rehgruppen auf, was vermutlich mit den auf der gleichen Höhe plazierten Fütterungsstellen im Zusammenhang steht. Im Nahbereich dieser Futterkrippen (100m Radius) halten sich signifikant mehr Rehgruppen auf. Die zusätzlich in diesen Gebieten sich aufhaltenden Tiere betreffen etwa 12.7% aller beobachteten Rehgruppen. Ab Distanzen über 100m entfernt von den Futterkrippen liessen sich keine Unterschiede im Vergleich zu einer gleichmässigen Verteilung nachweisen.
| Parameter | n | FG | Chi2 | p |
| Hangneigung | 115 | 6 | 29.9 | <0.001 |
| Höhe | 115 | 7 | 24.7 | <0.001 |
| Distanz Futterkrippe | 115 | 6 | 64.3 | <0.001 |
| Entwicklungsstadium | 115 | 5 | 14.0 | 0.015 |
| Anzahl Waldschichten | 115 | 2 | 0.96 | 0.62 |
| Hauptbaumart Oberschicht | 115 | 3 | 13.0 | 0.005 |
| Hauptbaumart Mittelschicht | 115 | 3 | 6.0 | 0.11 |
| Hauptbaumart Unterschicht | 115 | 3 | 1.6 | 0.67 |
| Anteil Nadelbäume Oberschicht | 115 | 4 | 13.2 | 0.01 |
| Anteil Nadelbäume Mittelschicht | 115 | 3 | 4.4 | 0.23 |
| Anteil Nadelbäume Unterschicht | 115 | 3 | 3.2 | 0.36 |
| Artenzahl Oberschicht | 115 | 4 | 9.8 | 0.04 |
| Artenzahl Mittelschicht | 115 | 4 | 8.9 | 0.06 |
| Artenzahl Unterschicht | 115 | 5 | 13.4 | 0.02 |
| Dichte Oberschicht | 115 | 5 | 8.8 | 0.12 |
| Dichte Mittelschicht | 112 | 4 | 6.5 | 0.16 |
| Dichte Unterschicht | 115 | 4 | 19.1 | <0.001 |
| Pflanzensoziologische Einheit | 115 | 9 | 63.3 | <0.001 |
Ein Unterschied zeigt sich auch in der Verteilung bezüglich Entwicklungsstadium (Alter) des Waldbestandes. Interessanterweise wurden in den ältesten Beständen (Stadien 6 und 7) weniger Rehgruppen als erwartet beobachtet. In den anderen Stadien zeigen sich keine signifikanten Unterschiede zum Angebot. Zwei im Sihlwald eng zusammenhängende Waldparameter sind Hauptbaumart und der Anteil Nadelbäume in der Oberschicht, weil Buche und Fichte zusammen flächenmässig rund 92% der Hauptbaumarten ausmachen. Bei der Hauptbaumart weisen Bestände mit Buche signifikant weniger, solche mit Fichte als Hauptbaumart signifikant mehr Rehgruppen als zu erwarten auf. Dem entsprechend sind signifikant weniger Rehgruppen in Beständen mit einem Anteil an Nadelbäumen von 0-10% beobachtet worden. Ebenfalls einen Unterschied zwischen Rehgruppenverteilung und Angebot gibt es bezüglich der Unterschichtdichte. Bestände mit 40-50% Dichte weisen signifikant mehr, solche mit 60-70% signifikant weniger Gruppen auf. Diese entgegengesetzte Reaktion zwischen benachbarten Dichteklassen muss vermutlich als Hinweis auf eine Überlagerung mit anderen Faktoren gedeutet werden. Ebenfalls in diesem Sinn ist vermutlich der signifikante Unterschied zwischen der Verteilung Rehgruppen und dem Angebot bezüglich der Anzahl Arten in der Unterschicht zu betrachten. In Gebieten mit nur einer Baumart in der Unterschicht waren signifikant mehr, in Gebieten mit drei Baumarten signifikant weniger Rehgruppen zu beobachten. Die anderen untersuchten Waldbestandesparameter zeigten keine signifikanten Unterschiede zwischen Rehgruppenverteilung und flächenmässigem Angebot. Beim Vergleich der Rehgruppenverteilung bezüglich der potentiellen pflanzensoziologischen Gesellschaft zeigte sich, dass in Gebieten mit zu erwartendem Waldmeister-Buchenwald signifikant mehr, in solchen mit Aronstab-Buchenwald signifikant weniger Rehgruppen anzutreffen waren.
Bei den hier verwendeten Bestandeserhebungen 1990 waren die Förster aufgefordert, die einzelnen Bestände danach zu beurteilen, ob diese als Reservatszonen ausgeschieden werden sollen. Interessanterweise ergab sich bei der Analyse, dass in den Gebieten, die als 'für eine Reservatszone geeignet' bezeichnet wurden, signifikant weniger Rehgruppen als erwartet beobachtet wurden (Chi2-Test, 2=11.1, n=109, FG=1, p<0.001).
B: Logistische Regression
Im Hinblick auf eine anschliessende Extrapolation auf den gesamten Sihlwald wurde als erstes Verfahren eine schrittweise logistische Regression (SLR) gerechnet. Das berechnete Modell ist hochsignifikant (Chi2=311.8, FG=48, p<0.0001) und erreicht eine überraschend hohe Zuordnungsgenauigkeit von fast 96%. Ins Modell wurden 9 Variablen aufgenommen (Tabelle 6), wovon die Höhenverteilung, Distanz zur Futterkrippe, Dichte der Unterschicht und Anteil Nadelbäume in der Oberschicht mit den Resultaten des vorhergehenden Kapitels übereinstimmen. Im Unterschied zu den vorangegangenen univariaten Vergleichen wurden zusätzlich folgende Variablen als signifikant miteinbezogen:
Von den in den univariaten Vergleichen signifikanten Variablen wurden jedoch die Hangneigung, Anzahl Baumarten in der Unterschicht und die potentielle Waldgesellschaft nicht ins Modell aufgenommen.
| Variable | Variable | B | S.E. | Wald | df | p | R | Exp(B) | Sig +/- |
| Dichte Oberschicht | 7.2256 | 3 | 0.065 | 0.0597 | |||||
| 20-40% | 20.1138 | 73.984 | 0.0739 | 1 | 0.7857 | 0 | 5.44E+08 | ||
| 40-60% | 8.872 | 3.4118 | 6.7619 | 1 | 0.0093 | 0.1177 | 7129.81 | - | |
| 60-80% | -1.4097 | 1.5277 | 0.8515 | 1 | 0.3561 | 0 | 0.2442 | ||
| Dichte Unterschicht | 13.5192 | 4 | 0.009 | 0.1267 | |||||
| 0-20% | 0.3016 | 3.5311 | 0.0073 | 1 | 0.9319 | 0 | 1.352 | ||
| 20-40% | 2.0667 | 2.4175 | 0.7308 | 1 | 0.3926 | 0 | 7.8985 | ||
| 40-60% | -9.1025 | 2.7611 | 10.8684 | 1 | 0.001 | -0.1606 | 0.0001 | + | |
| 60-80% | 3.2286 | 2.6084 | 1.5321 | 1 | 0.2158 | 0 | 25.2431 | ||
| Anzahl Waldschichten | 9.4015 | 2 | 0.0091 | 0.1253 | |||||
| 1 | -13.3414 | 4.9905 | 7.1468 | 1 | 0.0075 | -0.1223 | 0 | + | |
| 2 | 0.8388 | 1.9834 | 0.1788 | 1 | 0.6724 | 0 | 2.3135 | ||
| Distanz zur Futterkrippe | 0.0154 | 0.0041 | 14.0729 | 1 | 0.0002 | 0.1873 | 1.0155 | - | |
| Distanz zum Gewässer | -0.0759 | 0.0198 | 14.7045 | 1 | 0.0001 | -0.1922 | 0.9269 | + | |
| Hauptbaumart Unterschicht | 11.5847 | 3 | 0.009 | 0.1274 | |||||
| Ahorn | -15.9465 | 6.897 | 5.3458 | 1 | 0.0208 | -0.0986 | 0 | + | |
| Rotbuche | -17.0829 | 5.79 | 8.705 | 1 | 0.0032 | -0.1396 | 0 | + | |
| Esche | -11.6876 | 5.1715 | 5.1076 | 1 | 0.0238 | -0.095 | 0 | + | |
| Hauptbaumart Mittelschicht | 10.9339 | 4 | 0.0273 | 0.0923 | |||||
| Rotbuche | 13.6307 | 164.2967 | 0.0069 | 1 | 0.9339 | 0 | 831289.2 | ||
| - | 4.3883 | 164.314 | 0.0007 | 1 | 0.9787 | 0 | 80.4997 | ||
| Tanne | 13.2977 | 164.3072 | 0.0065 | 1 | 0.9355 | 0 | 595812.8 | ||
| Fichte | 17.4016 | 164.2967 | 0.0112 | 1 | 0.9156 | 0 | 36094461 | ||
| %Nadelbäume Oberschicht | 16.1065 | 4 | 0.0029 | 0.1535 | |||||
| 0-20% | -4.3491 | 2.3712 | 3.3639 | 1 | 0.0666 | -0.063 | 0.0129 | ||
| 20-40% | -16.3709 | 4.6364 | 12.4677 | 1 | 0.0004 | -0.1744 | 0 | + | |
| 40-60% | -21.045 | 5.9047 | 12.703 | 1 | 0.0004 | -0.1764 | 0 | + | |
| 60-80% | -7.0083 | 3.7821 | 3.4338 | 1 | 0.0639 | -0.0646 | 0.0009 | ||
| Höhe über Meer | 14.8944 | 7 | 0.0374 | 0.051 | |||||
| 470-510m | 10.6922 | 5.1327 | 4.3396 | 1 | 0.0372 | 0.0825 | 44009.15 | - | |
| 510-550m | 10.5611 | 4.8871 | 4.6699 | 1 | 0.0307 | 0.0881 | 38605.12 | - | |
| 550-590m | -6.9237 | 3.0574 | 5.1284 | 1 | 0.0235 | -0.0954 | 0.001 | + | |
| 590-630m | -5.3881 | 3.0262 | 3.1701 | 1 | 0.075 | -0.0583 | 0.0046 | ||
| 630-670m | -9.7484 | 4.8971 | 3.9627 | 1 | 0.0465 | -0.0755 | 0.0001 | + | |
| 670-710m | -1.1669 | 2.9033 | 0.1616 | 1 | 0.6877 | 0 | 0.3113 | ||
| 710-750m | -6.825 | 3.0183 | 5.113 | 1 | 0.0237 | -0.0951 | 0.0011 | + | |
| Konstante | 12.8306 | 164.5504 | 0.0061 | 1 | 0.9378 |
Die Variablen bzw. Klassen aus den Variablen, die sich signifikant negativ auf die Wahrscheinlichkeit des Rehvorkommens auswirken, sind eine grosse Distanz zur Futterkrippe, Dichte der Oberschicht = 40-60% und tief liegende Gebiete: 470-510m, 510-550m. Als signifikant positiv gewertet wurden: grosse Distanz zu Gewässern, Dichte der Unterschicht = 40-60%, Anteil Nadelbäume in der Oberschicht = 20-40% und 40-60%, Hauptbaumarten in der Unterschicht Ahorn, Buche und Esche, nur eine einzige Waldschicht vorkommend (d.h. nur Oberschicht) und die Höhenklassen 550-590m, 630-670m, 710-750m.
C: Schrittweise multiple (lineare) Regression
Als zweites Verfahren wurde eine multiple lineare Regression (MR) mit Dummyvariablen gerechnet. Dieses Modell ist ebenfalls hochsignifikant (Tabelle 7) und erklärt rund 77% der Variation.
| df | Sum of Squares | Mean Square | F | p | |
| Regression | 66 | 0.16639718 | 0.00252117 | 28.21 | 0.0001 |
| Error | 554 | 0.04950693 | 0.00008936 | ||
| Total | 620 | 0.21590411 |
Das Modell enthält mit 23 bedeutend mehr Variablen als die logistische Regression (Tabelle 8).
| Variable | Koeffizient | Error | Sum of Squares | F | p | Sig. +/- | |
| Konstante | 0.052809 | 0.0026746 | 0.03484 | 389.85 | 0.0001 | ||
| Distanz zum Gewässer | 0.000021 | 0.0000091 | 0.00046 | 5.18 | 0.0232 | + | |
| Distanz zur Futterkrippe | -0.000036 | 0.0000031 | 0.01210 | 135.36 | 0.0001 | - | |
| Hangneigung | 12-16 | -0.002370 | 0.0010651 | 0.00044 | 4.95 | 0.0265 | - |
| 16-20 | -0.003193 | 0.0013204 | 0.00052 | 5.85 | 0.0159 | - | |
| 20-24 | -0.004585 | 0.0017475 | 0.00062 | 6.89 | 0.0089 | - | |
| >24 | -0.002806 | 0.0014655 | 0.00033 | 3.67 | 0.0560 | ||
| Höhe über Meer | 510-550m | 0.006381 | 0.0017956 | 0.00113 | 12.63 | 0.0004 | + |
| 550-590m | 0.002486 | 0.0014799 | 0.00025 | 2.82 | 0.0935 | ||
| 630-670m | 0.003598 | 0.0012708 | 0.00072 | 8.02 | 0.0048 | + | |
| 750-790m | -0.006494 | 0.0018063 | 0.00115 | 12.92 | 0.0004 | - | |
| Dichte Unterschicht | 20-40% | -0.008499 | 0.0017855 | 0.00202 | 22.66 | 0.0001 | - |
| 40-60% | 0.008975 | 0.0015795 | 0.00289 | 32.29 | 0.0001 | + | |
| 80-100% | 0.007900 | 0.0026317 | 0.00081 | 9.01 | 0.0028 | + | |
| Dichte Oberschicht | 20-40% | 0.017804 | 0.0046793 | 0.00129 | 14.48 | 0.0002 | + |
| 60-80% | 0.012215 | 0.0014965 | 0.00595 | 66.62 | 0.0001 | + | |
| 100% | 0.009242 | 0.0019290 | 0.00205 | 22.95 | 0.0001 | + | |
| Dichte Mittelschicht | 40-60% | -0.004322 | 0.0022974 | 0.00032 | 3.54 | 0.0605 | |
| 60-80% | -0.055276 | 0.0132895 | 0.00155 | 17.30 | 0.0001 | - | |
| 80-100% | -0.049405 | 0.0089857 | 0.00270 | 30.23 | 0.0001 | - | |
| Artenzahl Unterschicht | 1 | 0.017665 | 0.0029226 | 0.00326 | 36.53 | 0.0001 | + |
| 2 | 0.008086 | 0.0028058 | 0.00074 | 8.31 | 0.0041 | + | |
| 3 | 0.005270 | 0.0026470 | 0.00035 | 3.96 | 0.0470 | + | |
| 4 | -0.006459 | 0.0026608 | 0.00053 | 5.89 | 0.0155 | - | |
| 5 | -0.006089 | 0.0027636 | 0.00043 | 4.85 | 0.0280 | - | |
| Artenzahl Oberschicht | 1 | 0.021735 | 0.0103944 | 0.00039 | 4.37 | 0.0370 | + |
| 2 | -0.018929 | 0.0030269 | 0.00349 | 39.11 | 0.0001 | - | |
| 3 | -0.020403 | 0.0019294 | 0.00999 | 111.83 | 0.0001 | - | |
| 4 | -0.009556 | 0.0018913 | 0.00228 | 25.53 | 0.0001 | - | |
| 6 | -0.010086 | 0.0021989 | 0.00188 | 21.04 | 0.0001 | - | |
| Artenzahl Mittelschicht | 2 | -0.016766 | 0.0018963 | 0.00699 | 78.17 | 0.0001 | - |
| 3 | -0.004040 | 0.0016793 | 0.00052 | 5.79 | 0.0165 | - | |
| 4 | -0.009317 | 0.0021911 | 0.00162 | 18.08 | 0.0001 | - | |
| 5 | -0.009870 | 0.0034180 | 0.00075 | 8.34 | 0.0040 | - | |
| 6 | -0.020955 | 0.0068891 | 0.00083 | 9.25 | 0.0025 | - | |
| %Nadelbäume Oberschicht | 20-40% | 0.006845 | 0.0019803 | 0.00107 | 11.95 | 0.0006 | + |
| 40-60% | 0.009963 | 0.0025391 | 0.00138 | 15.40 | 0.0001 | + | |
| 80-100% | 0.023368 | 0.0033188 | 0.00443 | 49.58 | 0.0001 | + | |
| 100% | -0.029489 | 0.0107646 | 0.00067 | 7.50 | 0.0064 | - | |
| %Nadelbäume Mittelschicht | 40-60% | 0.009115 | 0.0047203 | 0.00033 | 3.73 | 0.0540 | |
| 60-80% | 0.008093 | 0.0027093 | 0.00080 | 8.92 | 0.0029 | + | |
| 80-100% | -0.022035 | 0.0062559 | 0.00111 | 12.41 | 0.0005 | - | |
| Entwicklungsstufe | stufig | -0.013538 | 0.0029730 | 0.00185 | 20.74 | 0.0001 | - |
| 60-80 J. | -0.004489 | 0.0016887 | 0.00063 | 7.07 | 0.0081 | - | |
| 80-100 J. | -0.012214 | 0.0022087 | 0.00273 | 30.58 | 0.0001 | - | |
| 100-120 J. | -0.015014 | 0.0026355 | 0.00290 | 32.46 | 0.0001 | - | |
| > 120 J. | -0.009128 | 0.0037665 | 0.00052 | 5.87 | 0.0157 | - | |
| Exposition | 292.5-337.5 | -0.003970 | 0.0021686 | 0.00030 | 3.35 | 0.0677 | |
| Mittelschicht Esche | 20-40% | 0.031521 | 0.0067953 | 0.00192 | 21.52 | 0.0001 | + |
| 40-60% | 0.054695 | 0.0143077 | 0.00131 | 14.61 | 0.0001 | + | |
| Mittelschicht Tanne | 20-40% | 0.019598 | 0.0070023 | 0.00070 | 7.83 | 0.0053 | + |
| Oberschicht Buche | 40-60% | 0.008396 | 0.0014217 | 0.00312 | 34.88 | 0.0001 | + |
| 60-80% | 0.013469 | 0.0024918 | 0.00261 | 29.22 | 0.0001 | + | |
| 100% | -0.037676 | 0.0148759 | 0.00057 | 6.41 | 0.0116 | - | |
| Oberschicht Esche | 20-40% | -0.012429 | 0.0015257 | 0.00593 | 66.36 | 0.0001 | - |
| 40-60% | 0.012834 | 0.0032141 | 0.00142 | 15.95 | 0.0001 | + | |
| Oberschicht Ahorn | 20-40% | 0.010783 | 0.0042951 | 0.00056 | 6.30 | 0.0123 | + |
| Oberschicht Fichte | 20-40% | -0.006332 | 0.0020546 | 0.00085 | 9.50 | 0.0022 | - |
| 40-60% | -0.006916 | 0.0021559 | 0.00092 | 10.29 | 0.0014 | - | |
| 60-80% | -0.012688 | 0.0032583 | 0.00136 | 15.16 | 0.0001 | - | |
| Oberschicht Tanne | 80-100% | -0.046160 | 0.0141668 | 0.00095 | 10.62 | 0.0012 | - |
| Unterschicht Buche | 20-40% | -0.009223 | 0.0020389 | 0.00183 | 20.46 | 0.0001 | - |
| 40-60% | -0.014594 | 0.0021171 | 0.00425 | 47.52 | 0.0001 | - | |
| 60-80% | -0.028334 | 0.0030336 | 0.00780 | 87.24 | 0.0001 | - | |
| 80-100% | -0.011361 | 0.0047959 | 0.00050 | 5.61 | 0.0182 | - | |
| Unterschicht Esche | 20-40% | -0.008353 | 0.0018202 | 0.00188 | 21.06 | 0.0001 | - |
| 40-60% | -0.020687 | 0.0068905 | 0.00081 | 9.01 | 0.0028 | - |
Sechs der Variablen wurden schon in der logistischen Regression als signifikant ins Modell aufgenommen. Es sind dies Dichte der Ober- und Unterschicht, Distanz zur Futterkrippe und zum nächsten Gewässer, Nadelholzanteil in der Oberschicht und Höhe über Meer. Im Gegensatz dazu wurde die Anzahl Waldschichten nicht ins Modell aufgenommen. Andererseits wurden die folgenden Variablen ins multiple lineare Regressionsmodell aufgenommen, die sich bei der SLR nicht als signifikant erwiesen:
Neu hinzu kamen bei der multiplen Regression die berechneten Dichten einzelner Baumarten in den drei Waldschichten, die die Einteilung in Haupt- und Nebenbaumarten ersetzten.
Für den Vergleich der verwendeten Verfahren werden hier zuerst die selektierten Variablen und im anschliessenden Kapitel 4.3 auch die räumlichen Muster der Extrapolationen gegenübergestellt. In Tabelle 9 sind die Variablen aufgelistet, die in mindestens zwei Verfahren Signifikanzen aufwiesen.
| Chi2 | SLR | MR | |
| Dichte der Unterschicht | x | x | x |
| Distanz zur Futterkrippe | x | x | x |
| Nadelholzanteil der Oberschicht | x | x | x |
| Höhe über Meer | x | x | x |
| Dichte der Oberschicht | x | x | |
| Distanz zum Gewässer | . | x | x |
| Hangneigung | x | x | |
| Artenzahl der Ober-, Mittel- und Unterschicht | x | x | |
| Entwicklungsstadium | x | x | |
| Hauptbaumart der Oberschicht | x | x | . |
Vier Variablen wurden bei allen Verfahren als signifikant bewertet. Es sind dies Dichte der Unterschicht, Distanz zur Futterkrippe, Nadelholzanteil der Oberschicht und Höhe über Meer. Überall als günstig bewertet wurde eine kleine Distanz zur Futterkrippe, 40-60% Dichte der Unterschicht und Vorkommen von Nadelholz. Die Höhe über Meer als indirekte Variable ging unterschiedlich in die einzelnen Modelle ein. Weitere acht Variablen ergaben Signifikanzen in jeweils zwei Verfahren. Die restlichen Variablen gingen nur in jeweils eine der Analysemethoden als wichtig ein.
Mit Hilfe des berechneten logistischen Regressionsmodells (SLR) wurde nun im Geographischen Informationssystem eine Extrapolation auf den gesamten Sihlwald gerechnet. Das Ergebnis ist in Abbildung 8 dargestellt.
Wie zu erwarten zeigt sich eine gute Deckung der Rehbeobachtungen mit den Gebieten hoher Auftretenswahrscheinlichkeit von Rehen (hellgrüner Bereich). Es tritt ein Mosaikmuster von Flächen mit hoher und solchen mit niedriger Wahrscheinlichkeit hervor, bei dem die aufgrund der Beobachtungen erfolgten Einteilung in die drei Zentren (1) Schönenboden, (2) Biri-, Schröter- und Häuliboden und (3) Cholbenholz immer noch abgegrenzt werden können, wenn auch (1) und (3) grossflächiger als vermutet erscheinen (für Ortsbezeichnungen siehe Abbildung 2). Die erwähnte Lücke in den Beobachtungen im westlichen Mittelteil des Sihlwalds manifestiert sich vor allem in den höheren Lagen als grösseres Gebiet mit kleiner Wahrscheinlichkeit von Rehbeobachtungen. Ansonsten sind keine grosse zusammenhängende Teile mit kleiner Wahrscheinlichkeit erkennbar. Die Gesamtfläche der Gebiete mit einer Wahrscheinlichkeit von über 50% beträgt für den Sihlwald links der Sihl rund 56%. Wahrscheinlichkeiten über 5% ergeben sich bei 65% der Fläche. Die Gebiete der Stadtwaldungen rechts der Sihl und das Gebiet Winzelen nördlich der Albispassstrasse ergeben kleine Wahrscheinlichkeiten von Rehbeobachtungen. Die Zone mit einer Wahrscheinlichkeit über 50% entspricht in diesen Gebieten rund 26% der Fläche. Das entsprechende Flächenangebot für Auftretenswahrscheinlichkeiten über 5% beläuft sich auf 33% (Abbildung 8).
Die Extrapolation der Rehverteilung mittels multipler Regression (MR) auf den gesamten Sihlwald ist in Abbildung 9 dargestellt. Auffällig ist, dass rund 20% der Beobachtungspunkte in Gebieten liegen, in denen aufgrund der MR keine Rehe zu erwarten sind. Von den drei oben bezeichneten Zentren treten (1) Schönenboden und (2) Biri-, Schröter- und Häuliboden wieder deutlich hervor. Das Gebiet (3) Cholbenholz im Norden des Sihlwalds enthält in der Mitte eine grössere Fläche mit berechneten Rehdichten von 0 (Abbildung 9). Etwa 800m westlich davon weisen die steilen Hänge im Raum Schnabel ebenfalls kleine Werte (0) aus der MR auf. Als dritte grosse Fläche mit kleinen berechneten Dichtewerten tritt das Dreieck Spinnerweg-Waldmattstrasse/Weierbrunnen-strasse-Albishornstrasse hervor. Im Unterschied zur SLR weist jedoch ein ca. 100m breiter Streifen entlang der Krete zwischen Schnabelburg und Weierbrunnen Werte über 0 auf. Im südlichen Teil des Sihlwalds treten drei kleinere Flächen mit kleinen Werten hervor: Roregg, das Gebiet südlich des oberen Eschentobels und der Raum Egliboden. Letzterer enthält jedoch mindestens vier Rehbeobachtungen, so dass hier von der MR sicher eine Fehleinschätzung gemacht wird. Die Gesamtfläche der Gebiete mit berechneten Dichtewerten über 0 (kernel density estimation) beträgt für den Sihlwald links der Sihl rund 66%, in den Gebieten der Stadtwaldungen rechts der Sihl und nördlich der Albispassstrasse (Winzelen) rund 59%.
Um einen räumlichen Vergleich der beiden Modelle SLR und MR zu machen, wurden die Karten der Extrapolationen zu Positiv/Negativ-Karten (mit Rehvorkommen bzw. keine Rehe erwartet) wie folgt umgerechnet:
| Rehvorkommen erwartet | keine Rehe erwartet | |
| schrittweise logistische Regression: | Auftretenswahrscheinlichkeit 50% | Auftretenswahrscheinlichkeit < 50% |
| multiple lineare Regression: | Berechnete Dichte 0 | Berechnete Dichte > 0 |
Dann wurde eine Differenzkarte (Abbildung 10) berechnet, bei der nun Gebiete, die von beiden Modellen als positiv (mit erwartetem Rehvorkommen) bewertet wurden, grau eingefärbt sind, Gebiete, die von beiden Modellen als negativ (keine Rehe erwartet) bewertet wurden, schwarz markiert sind. Flächen, die unterschiedlich bewertet wurden, sind weiss dargestellt. Flächen kleiner als 1 Hektare wurden der umliegenden Fläche zugeteilt.
Die beiden Modelle stimmen in diesem recht groben Vergleich in
mehreren Punkten überein. Die schon mehrmals erwähnten
Gebiete (1) Schönenboden und (2) Biri-, Schröter- und
Häuliboden und die Lücke im Dreieck Spinnerweg-Waldmattstrasse/Weierbrunnenstrasse-Albishornstrasse
stimmen in der Bewertung überein. Die Gebiete Roregg und
Egliboden werden ebenfalls weitgehend übereinstimmend als
Flächen ohne erwartetem Rehvorkommen bewertet. Dagegen weist
der nördliche Sihlwald recht grosse Unterschiede zwischen
den Modellen auf. Beide Modelle stimmen jedoch im Gebiet Winzelen
nördlich des Albispasses wieder überein.