Die Austritte der Rehe aus dem Wald ins offene Kulturland zeigen zwei ausgeprägte Muster, die durch die beiden sich überlagernden Faktoren, Tageszeit und Jahreszeit, geformt werden. Zusätzlich existieren viele weniger deutliche Muster, von denen einige im folgenden ebenfalls dargestellt werden. Die Ergebnisse der Zählungen für Fuchs, Hase, Dachs und Mensch sind im Anhang zusammengestellt.
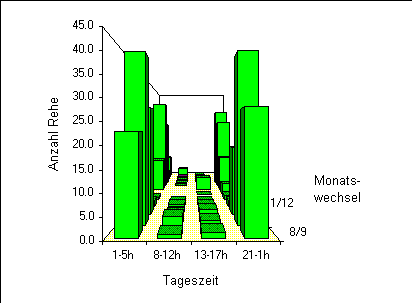
Ein stark ausgeprägtes Muster ist in der tageszeitlichen Nutzung der Waldränder ersichtlich (Abbildung 11).
Die Austritte aus dem Wald finden zu einem überwiegenden Teil - es handelt sich um über 95% der beobachteten Tiere - nachts statt.
Am Tag halten sich die Rehgruppen mit einer durchschnittlichen Distanz von 13m im Vergleich zur Nacht (51m) deutlich näher am Waldrand auf (MWU, n1=423, n2=18, z=5.41, p<0.0001). Zudem wurden liegende Rehe am Tag sehr selten beobachtet, wogegen in der Nacht rund 24% der Tiere lagen (Chi2=9.2, DF=1, n=981,p=0.003).
Es konnte kein Unterschied in der Anzahl beobachteter Rehe zwischen der ersten und der zweiten Nachthälfte festgestellt werden (Wilcoxon matched pairs signed rank Test, z=0.728, n=22, p=0.47). Ausserdem liessen sich weder Unterschiede bezüglich Distanz der Rehgruppen zum Waldrand (MWU, n1=226, n2=256, z=1.54, p=0.12) noch in der Gruppengrösse (MWU, n1=249, n2=267, z=0.81, p=0.42) zwischen den beiden Nachthälften nachweisen.
Auch am Tag zeigt sich kein Unterschied zwischen Morgen und Nachmittag bezüglich Anzahl beobachteter Rehe. Auf 36 Rundgängen wurden 16 Tiere am Morgen (8 Gruppen, 18 Rundgänge) und 16 Tiere am Nachmittag (10 Gruppen, 18 Rundgänge) gesehen.
Da die Austritte zu einem überwiegenden Teil nachts stattfinden, wird im folgenden auf die Austritte in der Nacht genauer eingegangen.
Das zweite neben dem circadianen sehr auffällige zeitliche Muster ist die saisonale Veränderung der Häufigkeit, mit der die Rehe aus dem Wald ins offene Kulturland austreten (Abbildung 12).
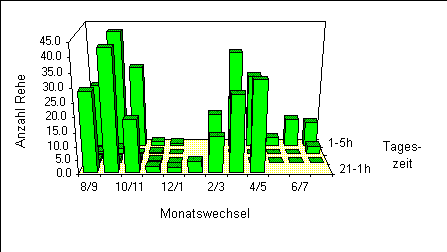
Dabei lassen sich grob vier etwa dreimonatige Phasen unterscheiden, die sich ungefähr nach den vier Jahreszeiten benennen lassen. Herbst und Frühling (8/9-10/11 bzw. 2/3-4/5) sind zwei Phasen mit sehr vielen Austritten, wobei das absolute Maximum im September/Oktober zu verzeichnen war. Im Winter (12/1-1/2) gab es im Vergleich dazu fast keine Austritte mehr. Abgesehen von der speziellen Situation im südlichsten Teil (vgl. Räumliche Verteilung) waren nur sehr sporadisch einzelne Tiere zu sehen. Im Sommer (5/6-7/8) wurden deutlich weniger Austritte als im Frühling und Herbst registriert.
Um zu prüfen, welche Monate sich anzahlmässig voneinander unterscheiden, wurden U-Tests der aufeinanderfolgenden Monate mit den einzelnen Rundgängen als Stichproben durchgeführt. Die Ergebnisse sind in Tabelle 10 dargestellt.
| Monatswechsel | n1 | n2 | z | p | |
| 8/9 | 9/10 | 6 | 6 | -2.40 | 0.016 |
| 9/10 | 10/11 | 6 | 6 | -2.72 | 0.007 |
| 10/11 | 11/12 | 6 | 6 | -2.90 | 0.004 |
| 11/12 | 12/1 | 6 | 6 | -0.75 | 0.454 |
| 12/1 | 1/2 | 6 | 1 | -1.56 | 0.120 |
| 1/2 | 2/3 | 1 | 2 | -1.23 | 0.221 |
| 2/3 | 3/4 | 2 | 6 | -2.01 | 0.044 |
| 3/4 | 4/5 | 6 | 6 | -0.64 | 0.520 |
| 4/5 | 5/6 | 6 | 3 | -2.38 | 0.017 |
| 5/6 | 6/7 | 3 | 3 | -2.09 | 0.037 |
| 6/7 | 7/8 | 3 | 3 | -0.44 | 0.658 |
| 7/8 | 8/9 | 3 | 6 | -1.81 | 0.071 |
Die in Abbildung 12 optisch in Erscheinung tretenden Unterschiede zwischen den Monaten lassen sich somit mit Ausnahme zwischen Jan/Feb (nur ein Rundgang!) und Feb/Mar statistisch nachweisen.
Bei den Beobachtungen am Tag scheinen häufiger Rehe in der
zweiten als in der ersten Jahreshälfte aus dem Wald herauskommen.
Zwischen Juli und Dezember wurden tendenziell auf mehr Rundgängen
Rehe beobachtet als zwischen Januar und Juni (Fischer Test, p=0.059,
n=38, zweiseitig).
Wie in Abbildung 13 ersichtlich ist, sind keine grossen Unterschiede in der Gruppengrösse im Verlauf des Jahres aufgetreten.
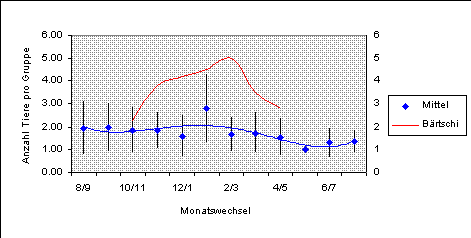
Statistisch lassen sich dennoch über die gesamte Untersuchungsdauer Unterschiede nachweisen (H-Test, DF=11, n=567, Chi2=36.32, p=0.0001). Zwischen aufeinanderfolgenden Monaten ergibt nur der Vergleich zwischen Mond 9 und 10 eine Irrtumswahrscheinlichkeit kleiner als 0.05 (MWU, n1=115, n2=9, z=2.29, p=0.022), was sich jedoch bei der nötigen Fehleradjustierung als nicht signifikant herausstellt (pkorrigiert=0.26).
Fasst man mehrere Monate zusammen, so zeigt sich, dass in der Zeit zwischen Apr/Mai bis Jul/Aug (Territorialzeit, Kurt 1991) die durchschnittliche Gruppengrösse mit 1.46 Rehen/Gruppe kleiner ist als im restlichen Jahr (1.86 Rehen/Gruppe; MWU, n1=166, n2=399, z=-4.99, p<0.00001).
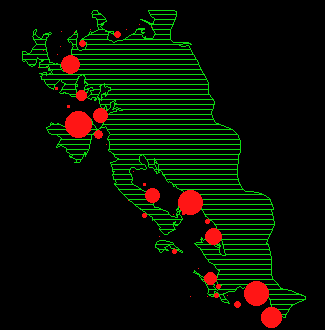
Im Verlauf des Untersuchungsjahres wurden auf fast allen Beobachtungsflächen mindestens einmal Rehe gesehen. Diejenigen Flächen, auf denen nie Rehe beobachtet wurden sind in Abbildung 3 als schwarze Felder dargestellt.
Es sind dies:
Diese Flächen scheinen keine auffälligen Merkmale gegenüber den von Rehen benutzten Gebieten aufzuweisen. Am erstaunlichsten ist, dass die grosse Fläche auf dem Albispass nicht genutzt wird, da dort regelmässig Hasen, Füchse und seltener auch Dachse auftraten. Die daran anschliessenden Wiesen wurden hingegen relativ intensiv genutzt.
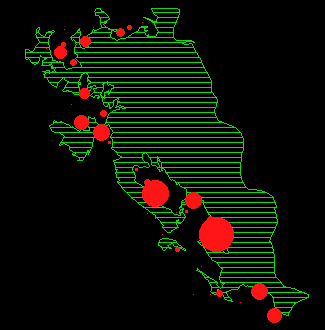
In Abbildung 14 ist die Rehverteilung für die ganze Beobachtungsperiode mittels einer 'Kernel Estimation' dargestellt. Die Flächen werden von den Rehen nicht gleichmässig genutzt, sondern es zeigen sich deutliche Nutzungszentren und daneben wenig genutzte Flächen. Im folgenden werden nun die jahreszeitlichen Veränderungen dieser Verteilung genauer untersucht.
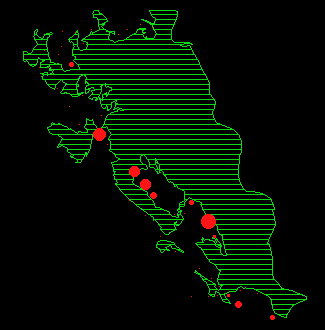
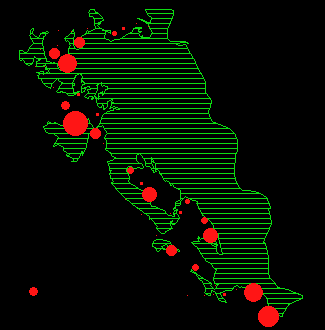
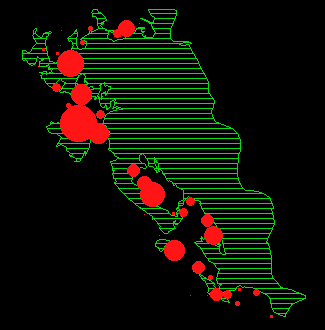
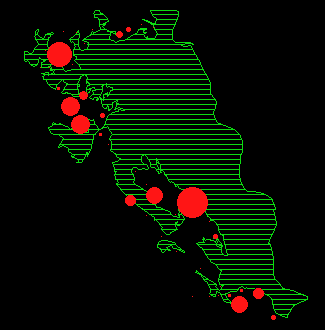
In Abbildung 15 ist für jeden Monat die durchschnittliche Anzahl Rehe pro Beobachtungspunkt dargestellt.
Während der meisten Zeit des Jahres verteilen sich die Austritte auf fast den gesamten Waldrand. Um Verteilungsunterschiede zwischen den einzelnen Monaten nachzuweisen, wurde ein Permutationsverfahren (MRPP, BIONDINI 1988) angewendet. Die Resultate sind in Tabelle 11 dargestellt.
| M.W. | 8/9 | 9/10 | 10/11 | 11/12 | 12/1 | 1/2 | 2/3 | 3/4 | 4/5 | 5/6 | 6/7 | |||||
| 9/10 | 0.00021 | |||||||||||||||
| 10/11 | 0.31512 | 0.00014 | ||||||||||||||
| 11/12 | 1.00000 | 0.29328 | 1.00000 | |||||||||||||
| 12/1 | 0.00038 | 0.09760 | 0.00007 | 0.06905 | ||||||||||||
| 1/2 | 0.07422 | 0.06708 | 0.00002 | 0.00016 | 0.00706 | |||||||||||
| 2/3 | 0.18140 | 0.02044 | 0.03755 | 0.01650 | 0.00043 | 0.00589 | ||||||||||
| 3/4 | 0.01638 | 0.10642 | 0.00373 | 0.52294 | 0.02131 | 0.10724 | 0.05752 | |||||||||
| 4/5 | 0.06871 | 0.02991 | 0.01258 | 0.94001 | 0.00936 | 0.11080 | 0.08111 | 0.52947 | ||||||||
| 5/6 | 1.00000 | 0.78604 | 1.00000 | 0.83468 | 0.24891 | 0.00035 | 0.03170 | 1.00000 | 1.00000 | |||||||
| 6/7 | 0.16579 | 0.02043 | 0.01921 | 0.00429 | 0.00043 | 0.00066 | 0.22408 | 0.05603 | 0.08037 | 0.01209 | ||||||
| 7/8 | 0.19531 | 0.02282 | 0.11664 | 0.02910 | 0.00050 | 0.00087 | 0.35961 | 0.06057 | 0.08643 | 0.05301 | 0.65936 | |||||
| p*0.05 (Bonferoni corrected(Cross&Chaffin)) = 0.001351 | ||||||||||||||||
| Monat | 8/9 | 9/10 | 10/11 | 11/12 | 12/1 | 1/2 | 2/3 | 3/4 | 4/5 | 5/6 | 6/7 | 7/8 | ||||
| n= | 156 | 259 | 111 | 16 | 8 | 14 | 30 | 188 | 177 | 9 | 30 | 29 | ||||
Bei näherer Betrachtung treten zwei Besonderheiten hervor:
In den Abbildungen 15 k und l zeigt sich noch eine weitere Besonderheit: die Austritte in diesen Sommermonaten (Jun/Jul und Jul/Aug) konzentrieren sich in den Gebieten Gom, Mittelalbis und Oberalbis, während sich die Austritte im Frühling und Herbst auf den ganzen Waldrand verteilen (Abbildung 15 a, b, c, h, i).
Die Höhenverteilung der Beobachtungen ist in Abbildung 16 dargestellt.
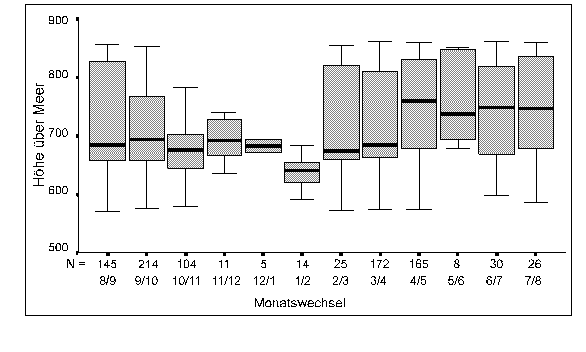
Obwohl das Untersuchungsgebiet nur eine geringe Höhenausdehnung aufweist, zeigen sich dennoch signifikante Unterschiede in der Höhenverteilung der Rehgruppen im Verlauf des Jahres (H-Test, n=531, Chi2=29.58, df=11, p=0.002). Im Spätherbst/Winter (Okt/Nov-Jan/Feb) sind die Austritte durchschnittlich in tieferen Lagen als in der übrigen Zeit (MWU, n1=71, n2=460, z=-3.90, p=0.0001). Eine Verschiebung der Höhenverteilung der Austritte scheint jedoch schon viel früher (Sep/Okt) zu beginnen (Abbildung 16).
In Bezug auf die Hangneigung lassen sich keine Unterschiede zwischen den einzelnen Monaten nachweisen (H-Test, n=532, Chi2=8.74, df=11, p=0.64). Beim Vergleich der Beobachtungen mit dem Angebot in den Beobachtungsflächen zeigen sich drei Abweichungen von den Erwartungswerten (Tabelle 12). In Gebieten mit einer Hangneigung von 12-16° wurden signifikant mehr, in den Hangneigungsklassen 4-8° und >24° weniger Rehe als erwartet beobachtet.
| Hangneigung | Beobachtet | Erwartet | Signifikant |
| 0-4 | 127 | 106.26 | - |
| 4-8 | 202 | 303.78 | sign. |
| 8-12 | 260 | 235.07 | - |
| 12-16 | 234 | 146.27 | sign. |
| 16-20 | 68 | 83.02 | - |
| 20-24 | 25 | 27.58 | - |
| 24-56 | 3 | 17.02 | sign. |
| Total | 919 | 919 |
Eine dritte sehr offensichtliche Komponente für die Verteilung der Rehe ist die Distanz zum Waldrand. Eine markante Distanzgrenze, die die Rehe nicht überschreiten würden, lässt sich innerhalb dieses Beobachtungsgebietes nicht feststellen. Im Vergleich zum Angebot zeigen sich jedoch drei Besonderheiten (Abbildung 17):
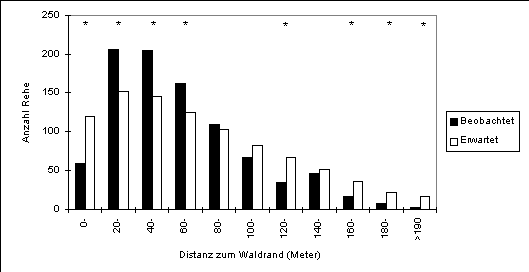
In Abbildung 18 sind die Aktivitäten der Rehe für die
vier Jahresabschnitte (s. 5.3) in die drei Gruppen 'liegen', 'äsen
& stehen' und 'diverse' zusammengefasst dargestellt. Die Anteile
der Aktivitätsgruppen in den vier Jahresabschnitten unterscheiden
sich signifikant (H-Test, Chi2=29.31, df=6, n=990,
p=0.00005).
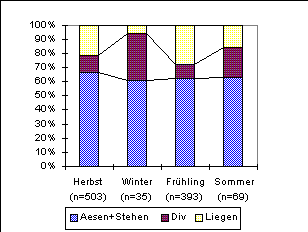
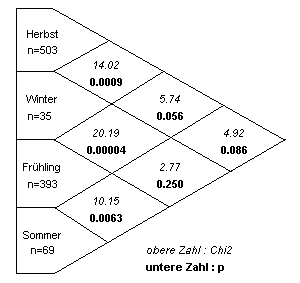
Vergleicht man nun die Zeitabschnitte einzeln, so lassen sich die Unterschiede zwischen Herbst-Winter, Winter-Frühling und Frühling-Sommer statistisch belegen. Die anderen paarweisen Vergleiche zeigen keine signifikanten Unterschiede (Tabelle 13). Liegende Rehe wurden im Winter seltener gesehen als in den anderen Jahreszeiten.
Interessanterweise lässt sich kein Unterschied zwischen liegenden
und äsenden Rehen bezüglich Distanz zum Waldrand nachweisen
(MWU, n1=376, n2=168, z=0.98, p=0.33),
hingegen zeigt sich bei liegenden Rehen eine etwas kleinere Hangneigung
(Median=8.6°) als bei äsenden (Median=10.3°; MWU,
n1=220, n2=429, z=3.96, p=0.0001).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}